Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Морфологія еукаріотичної клітиниСтр 1 из 3Следующая ⇒
Реферат З дисципліни”Біологія з основами генетики ” На тему: “Будова еукаріотичної клітини.Структурні компоненти цитоплазми” Виконав: Студент фармацевтичного Факультету(заочна форма навчання) Група 2А,1 курс Васильчук Володимир Валерійович Київ – 2016 Зміст 1. Морфологія еукаріотичної клітини
2. Гіпотези походження еукаріотів
2.1. Автогенетична гіпотеза 2.2. Ендосимбіотична гіпотеза 2.3. Синтетична гіпотеза
3. Первинні та вторинні ендосимбіози
4. Структурні компоненти цитоплазми.
5. Гіалоплазма, її склад і значення в метаболізмі клітини.
6. Органели, визначення і класифікація.
7. Мікро- і субмікроскопічна будова та функція органел загального призначення (мітохондрій, комплексу Гольджі, ендоплазматичної сітки, рибосом, полісом, центросом).
8. Органели спеціального призначення, їх види, локалізація і функції.
9. Включення: визначення, відмінності від органел, класифікація.
10. Морфофункціональна характеристика різних видів включень.
11. Загальна морфологія інтерфазних ядер
12.Література
Морфологія еукаріотичної клітини
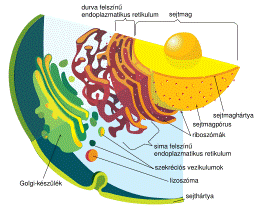
Мал.1 Клітина еукаріотів
Домен Я́дерні або Еукаріоти (Евкаріоти) (Eukaryota Whittaker & Margulis, 1978) — домен одно- та багатоклітинних організмів, що характеризуються переважно полігеномними клітинами, морфологічно сформованим ядром та наявністю мембранних субклітинних органел. Геноми еукаріотичної клітини представлені: а) ядерним геномом, зосередженим у ядрі і представленим ядерною ДНК; б) У більшості клітин мітохондріальним геномом, зосередженим у мітохондрії і представленим мітохондріальною ДНК; в) У деяких клітин пластидним (зокрема хлоропластним) геномом, що розташовується у пластиді, і представлений хлоропластною ДНК (генофором); г) геномом нуклеоморфу., який виявлений лише у кількох відділах водоростей у надзвичайно цікавій ядроподібній структурі, розташованій між оболонкою пластиди та особливою клітинною системою — хлоропластним ендоплазматичним ретикулумом. В нуклеоморфі виявлена власна, нуклеоморфна ДНК. Двогеномні клітини, в яких представлено ядерний та мітохондріальний геноми, характерні для грибів та тварин; тригеномні — з ядерним, мітохондріальним та пластидним геномами — для майже всіх рослин; чотиригеномні — з ядерним, мітохондріальним, пластидним та нуклеоморфними геномами, виявлені у хлорарахніофітових та криптофітових водоростей, а також у деяких видів з відділу Dinophyta.
Клітини прокаріотів (бактерій та архей) та еукаріотів на фенотипічному рівні схожі за наявністю ДНК та білок-синтезуючого апарату, представленого рибосомами; наявністю клітинних мембран; ферментних комплексів, що забезпечують процеси реплікації, транскрипції, трансляції та синтезу АТФ. До складу клітин прокаріотів та еукаріотів входять білки, жири, вуглеводи, нуклеїнові кислоти, мінеральні речовини та вода. Відміни у будові клітин прокаріотів та еукаріотів пов'язані, в першу чергу, з тим, що майже вся ДНК прокаріотів складається з однієї хромосоми, і у випадку бактерій не пов'язана з білками-гістонами (хоча навіть бактерії мають гістоноподібні білки). Еукаріоти, на відміну від прокаріотів, зазвичай не містять генів, організованих у великі оперони, процес цитокінезу (поділу ДНК) проходить за участю веретена поділу. Рибосоми у еукаріотів більші, ніж рибосоми прокаріотів (умовна вага прокаріотичних рибосом становить 70S, тоді як еукаріотичних — 80S). Багато клітин еукаріотів здатні до фаго- та піноцитозу, мають морфологічно оформлене ядро, часто також мітохондрії, пластиди, ендоплазматичний ретикулум, комплекс Гольджі, лізосоми, пероксисоми, мають іншу структуру джгутиків і джгутикових моторів, мають клітинний центр з центріолями. Для багатоьох клітин еукаріотів характерні мітоз, мейоз, повноцінний статевий процес, проте набагато нижча ймовірність горизонтального переносу генів. Зазвичай еукаріотичні клітини більші за розмірами, ніж прокаріотичні — їх середній діаметр становить біля 5-20 мікрон, тоді як у бактерії та архей — 0,5-2 мікрона. 2. Гіпотези походження еукаріотів За палеонтологічними даними, прокаріоти на нашій планеті виникли біля 3,2 млрд. років тому, тоді як еукаріоти є набагато молодшими — їх вік складає лише біля 1,6 млрд. років. Сьогодні в межах доменів бактерії та архей описано біля 40 тис. видів (хоча оцінки їх загальної кількості складають біля 10 млн. видів), тоді як у домені еукаріотів — біля 1,7 млн. видів.
Автогенетична гіпотеза Згідно з цією гіпотезою, евкаріотична клітина утворилась з прокаріотичної внаслідок розвитку плазмалемою системи інвагінацій, які надалі замкнулись навколо ділянок плазмалеми з ферментами дихального ланцюга, фотосинтетичними пігментами, нуклеоїда, відповідно утворивши мітохондрії, пластиди та ядро. Розвиток системи інвагінацій мембран призвів також до виникнення ендоплазматичної сітки, комплексу Гольджі, лізосом та ін. Проте автогенетична гіпотеза не змогла пояснити чимало фактів. Наприклад, чому мітохондрії та хлоропласти містять власну ДНК і розмножуються поділом, чому в мітохондріях та хлоропластах наявні легкі прокаріотичні рибосоми, чому поровий апарат мітохондрій та пластид подібний до порового апарату прокаріот, і відрізняється від порового апарату плазмалеми евкаріот, як виник мітоз та з'явились мікротрубочки, та ін. Ендосимбіотична гіпотеза Наприкінці ХІХ ст. видатний ботанік та фізіолог рослин М. С. Цвєт, на основі спостережень поділу хлоропласту та ядерця у зеленої водорості Spirogyra висловив припущення, що клітини рослин можливо представляють собою симбіотичний утвір з клітини-господаря та бактеріальних внутрішньоклітинних симбіонтів, які відомі як хлоропласти та ядерця. Це припущення у 20-40-х роках ХХ ст. було детально розроблено, в першу чергу, К. С. Мережковським у так звану ендосимбіотичну гіпотезу походження евкаріотичної клітини. Проте ідеї К. С. Мережковського не знайшли підтримки у широкого загалу біологів. Гіпотезу ендосимбіотичного походження евкаріот називали вражаючим витвором людської фантазії", «луна-парком», «непристойною темою у колі освічених біологів»[2]. У 60-х роках гіпотеза ендосимбіотичного походження евкаріотичної клітини була заново сформульована американською дослідницею Л. Маргеліс. Згідно з гіпотезою Мережковського-Маргеліс, евкаріотична клітина виникла внаслідок кількох ендосимбіозів: гіпотетична прокаріотична анаеробна клітина, здатна до фагоцитозу, захопила, проте не перетравила, а зберегла в цитоплазмі, аеробну гетеротрофну бактерію, яка трансформувалась в мітохондрію. Далі клітина-господар, що містила мітохондрію, вступила в симбіоз з рухливою спірохетоподібною гетеротрофною бактерією, яка дала початок джгутику. Після цього внаслідок автогенетичного процесу, пов'язаного з утворенням глибоких інвагінацій плазмалеми, навколо нуклеоїду клітини-господаря утворилось ядро. Далі базальні тіла джгутиків трансформувались в центри організації веретена поділу, внаслідок чого виник спочатку мітоз, а потім його модифікація — мейоз, разом із статевим процесом. Виникла перша гетеротрофна евкаріотична клітина. Нащадки цієї клітини, еволюція яких пішла по шляху вдосконалення фагоцитозу, дали початок тваринам. Ті ж нащадки, які еволюціонували в напрямку удосконалення осмотрофного живлення, дали початок грибам. Симбіоз гетеротрофної евкаріотичної клітини з синьозеленою водорістю призвів до появи рослин, оскільки симбіотична синьозелена водорість надалі трансформувалась у пластиду. Синтетична гіпотеза
Численні перевірки ендосимбіотичної гіпотези, виконані з широким використанням електронної мікроскопії, мікрохімічних та молекулярно-генетичних методів, з одного боку, підтвердили гіпотезу ендосимбіотичного походження мітохондрій та пластид, з іншого — автогенетичну гіпотезу походження ядра та більшості одномембранних клітинних органел. Таким чином, на початку ХХІ ст. теорія походження евкаріот сформувалась на основі синтезу обох гіпотез — як автогенетичної, так і ендосимбіотичної, і отримала назву синтетичної гіпотези походження евкаріот. Сьогодні вважають, що першим кроком на шляху виникнення евкаріот було утворення у гіпотетичного прокаріотичного пращура численних внутрішніх інвагінацій плазмалеми, які, з одного боку, замкнули прокаріотичний нуклеоїд у двомембранну оболонку (тобто утворили морфологічно оформлене ядро), а з іншого — призвели до утворення ендоплазматичної сітки та похідного від неї комплексу Гольджі, а також травних вакуолей та їх похідних — лізосом. На другому етапі, внаслідок горизонтального переносу гену, що кодує білок тубулін від спірохетоподібних бактерій, примітивні евкаріоти набули здатності до синтезу тубулінових мікротрубочок. В результаті у евкаріот з'явився цитоскелет, джгутики з базальним тілами, веретено поділу, мітоз. Базальні тіла джгутиків надалі у частини представників трансформувалися у клітинний центр, а порушення нормального мітозу (зокрема, скорочення інтерфази) призвели до виникнення мейозу і пов'язаного з ним статевого процесу. Група первинно безмітохондріальних евкаріот сьогодні розглядається як перше, найпримітивніше царство евкаріот — Hypochondria (Домітохондріальні, або Гіпохондріати). Всі гіпохондріати є одноклітинними гетеротрофами з тваринною стратегією живлення. На третьому етапі евкаріотична клітина утворила симбіотичний комплекс з прокаріотичною клітиною, схожою із сучасними альфа-протеобактеріями. Ця прокаріотична клітина надалі трансформувалась у мітохондрію. За результатами як цитологічних, так і молекулярних досліджень, найдавнішими мітохондріальними евкаріотами вважаються ті, які мають мітохондрії з особливими дископодібними кристами. Група мітохондріальних евкаріот з дископодібними кристами прийнята як таксон рангу царства — Discicristates (Дискокристати). В основі царства дискокристат знаходяться первинно гетеротрофні організми. На четвертому етапі евкаріоти розділилися на дві великі групи. Одна з цих груп має мітохондрії з трубчастими кристами, і складає царство Tubulocristates (Тубулокристати, трубчастокристні), друга — мітохондрії з переважно пластинчастими кристами, і приймається як царство Platycrsitates (платикристати, платівчастокристні). Подібно до дискокристат, в основі філ тубуло- та платикристат знаходяться первинно гетеротрофні організми із тваринною стратегією живлення.
На п'ятому етапі в евкаріотичному світі з'явилися перші рослини. За молекулярними та цитологічними даними, ця подія пов'язана з симбіозом гетеротрофної евкаріоти-платикристати з фотоавтотрофним прокаріотом — синьозеленою водорістю. Як наслідок цього симбіозу утворилась пластида, оточена двома мембранами, яка отримала назву первинно симбіотичної пластиди. Подальша дивергенція організмів з первинно симбіотичними пластидами обумовила виникнення у межах філи платикристат групи фотоавтотрофних відділів, які склали підцарство Plantae — рослини. Дивергенція гетеротрофних платикристат за типами живлення зумовила виникнення з одного боку, відділів з осмотрофним живленням — підцарства Fungi (гриби), з іншого — таксонів з фаготрофним живленням, що склали підцарство Animalia — тварини. Велика радіація тубулокристат відбувалась майже паралельно з платикристатами, проте була пов'язана не стільки з еволюцією за типами живлення, скільки з еволюцією клітинних покривів та джгутикового апарату. В основі тубулокристат залишились амебоїдні організми, які складають підцарство Ameboflagellates (амебо-флогеляти). Дві більш прогресивні філи представляють, з одного боку, таксони зі специфічними, альвеольованими покривами — Alveolates (альвеоляти), та з іншого — зі специфічними субмікроскопічними тричленними. Реферат З дисципліни”Біологія з основами генетики ” На тему: “Будова еукаріотичної клітини.Структурні компоненти цитоплазми” Виконав: Студент фармацевтичного Факультету(заочна форма навчання) Група 2А,1 курс Васильчук Володимир Валерійович Київ – 2016 Зміст 1. Морфологія еукаріотичної клітини
2. Гіпотези походження еукаріотів
2.1. Автогенетична гіпотеза 2.2. Ендосимбіотична гіпотеза 2.3. Синтетична гіпотеза
3. Первинні та вторинні ендосимбіози
4. Структурні компоненти цитоплазми.
5. Гіалоплазма, її склад і значення в метаболізмі клітини.
6. Органели, визначення і класифікація.
7. Мікро- і субмікроскопічна будова та функція органел загального призначення (мітохондрій, комплексу Гольджі, ендоплазматичної сітки, рибосом, полісом, центросом).
8. Органели спеціального призначення, їх види, локалізація і функції.
9. Включення: визначення, відмінності від органел, класифікація.
10. Морфофункціональна характеристика різних видів включень.
11. Загальна морфологія інтерфазних ядер
12.Література
Морфологія еукаріотичної клітини
Мал.1 Клітина еукаріотів
Домен Я́дерні або Еукаріоти (Евкаріоти) (Eukaryota Whittaker & Margulis, 1978) — домен одно- та багатоклітинних організмів, що характеризуються переважно полігеномними клітинами, морфологічно сформованим ядром та наявністю мембранних субклітинних органел. Геноми еукаріотичної клітини представлені: а) ядерним геномом, зосередженим у ядрі і представленим ядерною ДНК; б) У більшості клітин мітохондріальним геномом, зосередженим у мітохондрії і представленим мітохондріальною ДНК; в) У деяких клітин пластидним (зокрема хлоропластним) геномом, що розташовується у пластиді, і представлений хлоропластною ДНК (генофором); г) геномом нуклеоморфу., який виявлений лише у кількох відділах водоростей у надзвичайно цікавій ядроподібній структурі, розташованій між оболонкою пластиди та особливою клітинною системою — хлоропластним ендоплазматичним ретикулумом. В нуклеоморфі виявлена власна, нуклеоморфна ДНК. Двогеномні клітини, в яких представлено ядерний та мітохондріальний геноми, характерні для грибів та тварин; тригеномні — з ядерним, мітохондріальним та пластидним геномами — для майже всіх рослин; чотиригеномні — з ядерним, мітохондріальним, пластидним та нуклеоморфними геномами, виявлені у хлорарахніофітових та криптофітових водоростей, а також у деяких видів з відділу Dinophyta. Клітини прокаріотів (бактерій та архей) та еукаріотів на фенотипічному рівні схожі за наявністю ДНК та білок-синтезуючого апарату, представленого рибосомами; наявністю клітинних мембран; ферментних комплексів, що забезпечують процеси реплікації, транскрипції, трансляції та синтезу АТФ. До складу клітин прокаріотів та еукаріотів входять білки, жири, вуглеводи, нуклеїнові кислоти, мінеральні речовини та вода. Відміни у будові клітин прокаріотів та еукаріотів пов'язані, в першу чергу, з тим, що майже вся ДНК прокаріотів складається з однієї хромосоми, і у випадку бактерій не пов'язана з білками-гістонами (хоча навіть бактерії мають гістоноподібні білки). Еукаріоти, на відміну від прокаріотів, зазвичай не містять генів, організованих у великі оперони, процес цитокінезу (поділу ДНК) проходить за участю веретена поділу. Рибосоми у еукаріотів більші, ніж рибосоми прокаріотів (умовна вага прокаріотичних рибосом становить 70S, тоді як еукаріотичних — 80S). Багато клітин еукаріотів здатні до фаго- та піноцитозу, мають морфологічно оформлене ядро, часто також мітохондрії, пластиди, ендоплазматичний ретикулум, комплекс Гольджі, лізосоми, пероксисоми, мають іншу структуру джгутиків і джгутикових моторів, мають клітинний центр з центріолями. Для багатоьох клітин еукаріотів характерні мітоз, мейоз, повноцінний статевий процес, проте набагато нижча ймовірність горизонтального переносу генів. Зазвичай еукаріотичні клітини більші за розмірами, ніж прокаріотичні — їх середній діаметр становить біля 5-20 мікрон, тоді як у бактерії та архей — 0,5-2 мікрона. 2. Гіпотези походження еукаріотів За палеонтологічними даними, прокаріоти на нашій планеті виникли біля 3,2 млрд. років тому, тоді як еукаріоти є набагато молодшими — їх вік складає лише біля 1,6 млрд. років. Сьогодні в межах доменів бактерії та архей описано біля 40 тис. видів (хоча оцінки їх загальної кількості складають біля 10 млн. видів), тоді як у домені еукаріотів — біля 1,7 млн. видів. Автогенетична гіпотеза Згідно з цією гіпотезою, евкаріотична клітина утворилась з прокаріотичної внаслідок розвитку плазмалемою системи інвагінацій, які надалі замкнулись навколо ділянок плазмалеми з ферментами дихального ланцюга, фотосинтетичними пігментами, нуклеоїда, відповідно утворивши мітохондрії, пластиди та ядро. Розвиток системи інвагінацій мембран призвів також до виникнення ендоплазматичної сітки, комплексу Гольджі, лізосом та ін. Проте автогенетична гіпотеза не змогла пояснити чимало фактів. Наприклад, чому мітохондрії та хлоропласти містять власну ДНК і розмножуються поділом, чому в мітохондріях та хлоропластах наявні легкі прокаріотичні рибосоми, чому поровий апарат мітохондрій та пластид подібний до порового апарату прокаріот, і відрізняється від порового апарату плазмалеми евкаріот, як виник мітоз та з'явились мікротрубочки, та ін. Ендосимбіотична гіпотеза Наприкінці ХІХ ст. видатний ботанік та фізіолог рослин М. С. Цвєт, на основі спостережень поділу хлоропласту та ядерця у зеленої водорості Spirogyra висловив припущення, що клітини рослин можливо представляють собою симбіотичний утвір з клітини-господаря та бактеріальних внутрішньоклітинних симбіонтів, які відомі як хлоропласти та ядерця. Це припущення у 20-40-х роках ХХ ст. було детально розроблено, в першу чергу, К. С. Мережковським у так звану ендосимбіотичну гіпотезу походження евкаріотичної клітини. Проте ідеї К. С. Мережковського не знайшли підтримки у широкого загалу біологів. Гіпотезу ендосимбіотичного походження евкаріот називали вражаючим витвором людської фантазії", «луна-парком», «непристойною темою у колі освічених біологів»[2]. У 60-х роках гіпотеза ендосимбіотичного походження евкаріотичної клітини була заново сформульована американською дослідницею Л. Маргеліс. Згідно з гіпотезою Мережковського-Маргеліс, евкаріотична клітина виникла внаслідок кількох ендосимбіозів: гіпотетична прокаріотична анаеробна клітина, здатна до фагоцитозу, захопила, проте не перетравила, а зберегла в цитоплазмі, аеробну гетеротрофну бактерію, яка трансформувалась в мітохондрію. Далі клітина-господар, що містила мітохондрію, вступила в симбіоз з рухливою спірохетоподібною гетеротрофною бактерією, яка дала початок джгутику. Після цього внаслідок автогенетичного процесу, пов'язаного з утворенням глибоких інвагінацій плазмалеми, навколо нуклеоїду клітини-господаря утворилось ядро. Далі базальні тіла джгутиків трансформувались в центри організації веретена поділу, внаслідок чого виник спочатку мітоз, а потім його модифікація — мейоз, разом із статевим процесом. Виникла перша гетеротрофна евкаріотична клітина. Нащадки цієї клітини, еволюція яких пішла по шляху вдосконалення фагоцитозу, дали початок тваринам. Ті ж нащадки, які еволюціонували в напрямку удосконалення осмотрофного живлення, дали початок грибам. Симбіоз гетеротрофної евкаріотичної клітини з синьозеленою водорістю призвів до появи рослин, оскільки симбіотична синьозелена водорість надалі трансформувалась у пластиду. Синтетична гіпотеза Численні перевірки ендосимбіотичної гіпотези, виконані з широким використанням електронної мікроскопії, мікрохімічних та молекулярно-генетичних методів, з одного боку, підтвердили гіпотезу ендосимбіотичного походження мітохондрій та пластид, з іншого — автогенетичну гіпотезу походження ядра та більшості одномембранних клітинних органел. Таким чином, на початку ХХІ ст. теорія походження евкаріот сформувалась на основі синтезу обох гіпотез — як автогенетичної, так і ендосимбіотичної, і отримала назву синтетичної гіпотези походження евкаріот. Сьогодні вважають, що першим кроком на шляху виникнення евкаріот було утворення у гіпотетичного прокаріотичного пращура численних внутрішніх інвагінацій плазмалеми, які, з одного боку, замкнули прокаріотичний нуклеоїд у двомембранну оболонку (тобто утворили морфологічно оформлене ядро), а з іншого — призвели до утворення ендоплазматичної сітки та похідного від неї комплексу Гольджі, а також травних вакуолей та їх похідних — лізосом. На другому етапі, внаслідок горизонтального переносу гену, що кодує білок тубулін від спірохетоподібних бактерій, примітивні евкаріоти набули здатності до синтезу тубулінових мікротрубочок. В результаті у евкаріот з'явився цитоскелет, джгутики з базальним тілами, веретено поділу, мітоз. Базальні тіла джгутиків надалі у частини представників трансформувалися у клітинний центр, а порушення нормального мітозу (зокрема, скорочення інтерфази) призвели до виникнення мейозу і пов'язаного з ним статевого процесу. Група первинно безмітохондріальних евкаріот сьогодні розглядається як перше, найпримітивніше царство евкаріот — Hypochondria (Домітохондріальні, або Гіпохондріати). Всі гіпохондріати є одноклітинними гетеротрофами з тваринною стратегією живлення. На третьому етапі евкаріотична клітина утворила симбіотичний комплекс з прокаріотичною клітиною, схожою із сучасними альфа-протеобактеріями. Ця прокаріотична клітина надалі трансформувалась у мітохондрію. За результатами як цитологічних, так і молекулярних досліджень, найдавнішими мітохондріальними евкаріотами вважаються ті, які мають мітохондрії з особливими дископодібними кристами. Група мітохондріальних евкаріот з дископодібними кристами прийнята як таксон рангу царства — Discicristates (Дискокристати). В основі царства дискокристат знаходяться первинно гетеротрофні організми. На четвертому етапі евкаріоти розділилися на дві великі групи. Одна з цих груп має мітохондрії з трубчастими кристами, і складає царство Tubulocristates (Тубулокристати, трубчастокристні), друга — мітохондрії з переважно пластинчастими кристами, і приймається як царство Platycrsitates (платикристати, платівчастокристні). Подібно до дискокристат, в основі філ тубуло- та платикристат знаходяться первинно гетеротрофні організми із тваринною стратегією живлення. На п'ятому етапі в евкаріотичному світі з'явилися перші рослини. За молекулярними та цитологічними даними, ця подія пов'язана з симбіозом гетеротрофної евкаріоти-платикристати з фотоавтотрофним прокаріотом — синьозеленою водорістю. Як наслідок цього симбіозу утворилась пластида, оточена двома мембранами, яка отримала назву первинно симбіотичної пластиди. Подальша дивергенція організмів з первинно симбіотичними пластидами обумовила виникнення у межах філи платикристат групи фотоавтотрофних відділів, які склали підцарство Plantae — рослини. Дивергенція гетеротрофних платикристат за типами живлення зумовила виникнення з одного боку, відділів з осмотрофним живленням — підцарства Fungi (гриби), з іншого — таксонів з фаготрофним живленням, що склали підцарство Animalia — тварини. Велика радіація тубулокристат відбувалась майже паралельно з платикристатами, проте була пов'язана не стільки з еволюцією за типами живлення, скільки з еволюцією клітинних покривів та джгутикового апарату. В основі тубулокристат залишились амебоїдні організми, які складають підцарство Ameboflagellates (амебо-флогеляти). Дві більш прогресивні філи представляють, з одного боку, таксони зі специфічними, альвеольованими покривами — Alveolates (альвеоляти), та з іншого — зі специфічними субмікроскопічними тричленними.
|
|||||||||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 258; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.144.102.239 (0.03 с.) |