Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Некоторые физические свойства и параметры мембран
С появлением электронного микроскопа (см. § 23.2) впервые открылась возможность познакомиться со строением мембран. Тогда обнаружилось, что плазматическая мембрана животных и растительных клеток выглядит как трехслойная структура. На рис. 11.7 изображена электронная микрофотография плазматической мембраны эритроцита. Видно, что мембрана состоит из светлого слоя, соответствующего фосфолипидам бислоя, и двух темных слоев — они представляют собой полярные головки и белки. Толщина мембран в зависимости от вида составляет величину от 4 до 13 нм. Измерение подвижности молекул мембран и диффузии частиц через мембрану свидетельствует о том, что билипидный слой ведет себя подобно жидкости. В то же время мембрана является упорядоченной структурой. Эти два фактора заставляют думать, что липиды в мембране при ее естественном функционировании находятся в жидкокристаллическом состоянии (см. § 8.2).
Вязкость липидного бислоя на два порядка больше вязкости воды и соот-
В жидкокристаллическом состоянии отдельная жирнокислотная цепь может принимать много различных конфигураций из-за вращения вокруг С—С связей. При том возможно образование в бислое полостей — «кинков» (от англ. kink — петля). В этих полостях могут находиться различные молекулы, захваченные из дространства вне мембраны. При тепловом движении хвостов липидов происходит движение такого «кинка», а вместе с ним и молекул поперек мембраны или вдоль нее (рис. 11.9). Проницаемость мембран для различных веществ зависит от поверхностного заряда, который создается заряженными головками липидов, придающими мембране преимущественно отрицательный заряд. Это приводит к тому, что на границе мембрана — вода создается межфазный скачок потенциала (поверхностный потенциал) того же знака, что и заряд на мембране. Величина этого потенциала играет большую роль в процессах связывания ионов мембраной. Помимо поверхностного потенциала, для нормального функционирования ферментных и рецепторных мембранных комплексов огромное значение имеет трансмембранный потенциал, природа которого будет рассмотрена ниже. Величина этого потенциала составляет 60—90 мВ (со знаком минус со стороны цитоплазмы). Из-за очень малой толщины мембран напряженность электрического поля в них достигает величины около (6—9) • 106 В/м.
Мембрана по своей структуре напоминает плоский конденсатор, обкладки которого образованы поверхностными белками, а роль диэлектрика выполняет липидный бислой. Емкость такого конденсатора составляет значительную величину (табл. 18). Используя формулу плоского конденсатора, можно оценить диэлектрическую проницаемость е гидрофобной и гидрофильной областей мембран, зная пределы изменения толщины мембраны. Такие оценки дают для фосфолипидной области мембраны значение ε = 2,0—2,2, а для гидрофильной части ε = 10—20. В табл. 18 приведены некоторые физические параметры биологических мембран и в сравнении с ними — те же параметры для искусственно приготовленных липидных бислоев. Таблица 18. Физические свойства биологических мембран и липидных бислоев
Мембраны обладают высокой прочностью на разрыв, устойчивостью и гибкостью. По электроизоляционным свойствам они значительно превосходят многие изоляционные материалы, применяемые в технике. Общая площадь мембран в органах и тканях достигает огромных размеров. Так, суммарная площадь клеточных мембран печени крысы, весящей всего 6 г, составляет несколько сотен квадратных метров. Клетки, как правило, имеют микроскопические размеры, поэтому отношение их поверхности к объему очень велико. Благодаря этому клетки располагают достаточной площадью для обеспечения многочисленных процессов, протекающих на мембранах. Одним из наиболее важных из них является процесс переноса веществ из клетки и в клетку.
§ 11.3. Перенос молекул (атомов) через мембраны.Уравнение Фика Важным элементом функционирования мембран является их способность пропускать или не пропускать молекулы (атомы) и ионы. Существенно, что вероятность такого проникновения частиц зависит как от направления их перемещения, например в клетку или из клетки, так и от разновидности молекул и ионов. Эти вопросы рассматриваются в разделе физики, относящемся - к явлениям переноса. Таким термином называют необратимые F, процессы, в результате которых в физической системе происходит пространственное перемещение (перенос) массы, импульса, энергии, заряда или какой-либо другой физической величины. К явлениям переноса относят диффузию (перенос массы вещества), вязкость (перенос импульса), теплопроводность (перенос энергии), электропроводность (перенос электрического заряда). Здесь и в следующих параграфах рассматриваются наиболее существенные для биологических мембран явления: перенос вещества и перенос заряда. Как синоним переноса частиц в биофизике широкое распространение получил также термин транспорт частиц. Выведем основное уравнение диффузии (уравнение Фика), рассматривая процесс переноса в жидкостях. Пусть через некоторую площадку S (рис. 11.10) во всех направлениях перемещаются молекулы жидкости. Учитывая теорию: молекулярного строения жидкости (см. § 7.6), можно сказать, что молекулы пересекают площадку, перескакивая из одного положения равновесия в другое.
выделенных параллелепипедов имеется Sin молекул. Предположим, что концентрация молекул изменяется в пространстве, в левом (1) выделенном параллелепипеде концентрация равна n1 а в правом (2) — п2. Следовательно, в одном параллелепипеде SZnx молекул, а в другом - Sln2 молекул. Все молекулы вследствие хаотичного их движения можно условно представить шестью группами, каждая из которых перемещается вдоль или против направления одной из осей координат. Отсюда следует, что в направлении, перпендикулярном площадке S, вдоль оси ОХот первого параллелепипеда перескакивает 1/6 Sln1 молекул, а противоположно оси ОХ от второго параллелепипеда перескакивает 1/6 Sln2 молекул.
Время Δt «пролета» этими молекулами площадки S может быть найдено следующим образом. Предположим, что все молекулы из выделенных объемов движутся с одинаковыми средними скоростями <v>. Тогда молекулы в объеме 1 или 2, дошедшие до площадки S, пересекают ее в течение промежутка времени
Подставляя в ( 11.1)выражение для средней скорости ύ из (7.20), получаем Δt=(l/δ)*τ (11.2)
где т — среднее время «оседлой жизни» молекулы, оно может рассматриваться как среднее время перескока. «Баланс» переноса молекул через площадку S за промежуток времени Δ t равен
Умножая (11.3)на массу т отдельной молекулы и деля на Δ t, находим массовый поток сквозь площадку S:
т. е. масса вещества, которая за 1 с переносится через площадку S. Изменение концентрации п2 - п1 молекул можно представить как произведение dn/dxна расстояние 2δ между выделенными объемами:
В уравнении (11.4) заменим Δ t согласно (11.2)и (п2 - п1) согласно (11.5): Отношение потока к площади S, через которую он переноситься, называется плотностью потока:
Произведение массы молекулы на их концентрацию есть плотность вещества (парциальная плотность):
Это есть уравнение диффузии (уравнение Фика), которое обычно записывают в виде:
Знак «-» показывает, что суммарная плотность потока вещества при диффузии направлена в сторону уменьшения плотности (в сторону, противоположную градиенту плотности), D — коэффициент диффузии, применительно к рассмотренному примеру диффузии в жидкости он равен
Как видно из ( 11.10),единица измерения коэффициента диффузии [м2/с]. Уравнение диффузии можно записать не только для плотности
А. Эйнштейн показал, что коэффициент диффузии пропорционален температуре:
И поэтому вместо (11.11) имеем
В формуле (11.12) и далее ит — подвижность диффундирующих молекул (частиц), выраженная для моля. Вообще говоря, подвижностью диффундирующей частицы (молекулы, атома, иона, электрона) и называют коэффициент пропорциональности между скоростью v частицы и силой f, двигающей частицу, в том случае, когда на частицу не действуют другие силы (например, трение или соударение с другими частицами) и она перемещается равномерно:
Как видно из (11.14), единица подвижности 1 м/(с • Н). Величины um и u связаны через постоянную Авогадро:
Преобразуем уравнение (11.9) применительно к биологической мембране. Будем считать, что концентрация частиц, диффундирующих через мембрану, изменяется в мембране по линейному закону (рис. 11.11). Молярные концентрации частиц внутри и вне клетки соответственно равны ci и с0. Молярная концентрация этих же частиц в мембране изменяется от внутренней к наружной ее части соответственно от сmi до см0. Учитывая линейное изменение концентрации молекул, запишем
где I — толщина мембраны, тогда вместо (11.11) имеем
Практически доступнее определить молярные концентрации частиц не внутри мембраны (cMi и см0), а вне мембраны: в клетке (сi) и снаружи клетки (с0). Считают, что отношение граничных значенийконцентраций в мембране равно отношению концентраций в прилегающих к мембране слоях: см0/сMi = co/ci, откуда
где k — коэффициент распределения вещества (частиц) между мембраной и окружающей средой (обычно водная фаза). Из (11.18) следует
Подставляя (11.19) в (11.17), имеем
Пусть
где Р — коэффициент проницаемости. В результате получаем уравнение для плотности потока вещества при диффузии через биологическую мембрану:
Уравнение Нернста—Планка. Перенос ионов через мембраны
Как известно, на мембране существует разность потенциалов, следовательно, в мембране имеется электрическое поле. Оно оказывает влияние на диффузию заряженных частиц (ионов и электронов). Между напряженностью поля Е и градиентом потенциала dφ/dx существует известное соотношение (см. § 12.1):
Заряд иона равен Ze. На один ион действует сила f=Ze(dφ/dx); сила, действующая на 1 моль ионов, равна где F — постоянная Фарадея, F = eNA.
 Скорость направленного движения ионов пропорциональна действующей силе [см. (11.4), (11.5)]: Скорость направленного движения ионов пропорциональна действующей силе [см. (11.4), (11.5)]:
Чтобы найти поток вещества (ионов), выделим объем электролита (рис. 11.12) в виде прямоугольного параллелепипеда с ребром, численно равным скорости ионов. Все ионы, находящиеся в параллелепипеде, за 1 с пройдут через площадку S. Это и будет поток Ф. Число молей этих ионов
можно найти, умножая объем параллелепипеда (vS) на молярную концентрацию ионов с:
Плотность потока вещества найдем, используя формулы (11.24) и (11.25):
В общем случае перенос ионов определяется двумя факторами: неравномерностью их распределения, т. е. градиентом концентрации [см. (11.11)], и воздействием электрического поля [см. (11.26)]: Это уравнение Нернста — Планка. Используя выражение для подвижности (11.12), преобразуем уравнение (11.27) к виду
Это другая форма записи уравнения Нернста—Планка
Используем уравнение Нернста—Планка для установления зависимости плотности диффузионного потока от концентрации ионов и от напряженности электрического поля. Предположим, система находится в стационарном состоянии, т. е. плотность потока Jпостоянна. Электрическое поле в мембране примем за однородное, следовательно, напряженность поля одинакова, а потенциал линейно изменяется с расстоянием. Это позволит считать, что
Упростим запись слагаемого в уравнении (11.28):
где — — —
вспомогательная величина (безразмерный потенциал). С учетом (11.29) получим уравнение Нернста—Планка в виде
Разделим переменные и проинтегрируем уравнение:
Потенцируя (11.31), получаем
откуда
Преобразуем формулу (11.32), учитывая выражения (11.19) и (11.20):
Вообще говоря, формула (11.33) справедлива как для положительных (Z > 0, у > 0), так и для отрицательных {Z < 0, ψ < 0) ионов. Однако для отрицательных ионов целесообразно видоизменить это выражение, подставив в него отрицательное значение безразмерного потенциала:
Разделим числитель и знаменатель этого выражения на е-ψ: При использовании этой формулы необходимо помнить, что отрицательные значения Z и ψ уже учтены в самой формуле, т. е. ψ— положительная величина. Уравнения (11.33) и (11.34) устанавливают связь плотности стационарного потока ионов с тремя величинами: 1) проницаемостью мембран для данного иона, которая характеризует взаимодействие мембранных структур с ионом; 2) электрическим полем; 3) молярной концентрацией ионов в водном растворе, окружающем мембрану {сi и с0). Проанализируем частные случаи уравнения (11.33):
а) ψ = 0, что означает либо Z = 0 (нейтральные частицы), либо отсутствие электрического поля в мембране (φм = 0), либо и то, и другое:
Найдем пределы отдельных сомножителей. 1.
2.Отсюда получаем, как и следовало ожидать, уравнение (11.21):
б) одинаковая молярная концентрация ионов по разные стороны от мембраны (ci = с0 = с) при наличии электрического поля:
Это соответствует электропроводимости в электролите (см. § 12.9). Для нейтральных частиц (Z = 0 и ψ=0) J = 0; в) если мембрана непроницаема для частиц (Р = 0), то, естественно, плотность потока равна нулю.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 636; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.133.109.30 (0.046 с.) |

 ветствует приблизительно вязкости растительного масла. Однако при понижении температуры происходит фазовый переход, в результате которого липиды бислоя превращаются в гель (твердо-кристаллическое состояние). На рис. 8 схематически представлен процесс «плавления» мембранных фосфолипидов при увеличении температуры (слева направо). Очевидно, что при этом меняется толщина двойного слоя — в состоянии геля (рис. 11.8, а) она больше, чем в жидкокристаллическом (рис. 11.8, б). При фазовых переходах в бислое могут образовываться каналы, по которым через мембрану способны проходить различные ионы и низкомолекулярные соединения, размер которых не превышает 1—3 нм.
ветствует приблизительно вязкости растительного масла. Однако при понижении температуры происходит фазовый переход, в результате которого липиды бислоя превращаются в гель (твердо-кристаллическое состояние). На рис. 8 схематически представлен процесс «плавления» мембранных фосфолипидов при увеличении температуры (слева направо). Очевидно, что при этом меняется толщина двойного слоя — в состоянии геля (рис. 11.8, а) она больше, чем в жидкокристаллическом (рис. 11.8, б). При фазовых переходах в бислое могут образовываться каналы, по которым через мембрану способны проходить различные ионы и низкомолекулярные соединения, размер которых не превышает 1—3 нм.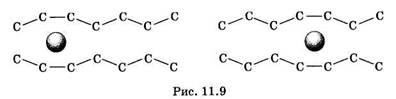
 На расстояниях, равных среднему перемещению 5 молекул (среднее расстояние между молекулами жидкости), вправо и влево от площадки построим прямоугольные параллелепипеды небольшой толщины l (l <<δ). Объем каждого параллелепипеда равен Sl. Если п — концентрация молекул, то внутри
На расстояниях, равных среднему перемещению 5 молекул (среднее расстояние между молекулами жидкости), вправо и влево от площадки построим прямоугольные параллелепипеды небольшой толщины l (l <<δ). Объем каждого параллелепипеда равен Sl. Если п — концентрация молекул, то внутри





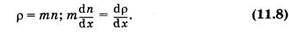

 или градиент молярной концентрации
или градиент молярной концентрации 

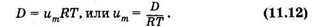

















 , где φм — разность потенциалов на мембране.
, где φм — разность потенциалов на мембране.









 Эту неопределенность можно раскрыть по правилу Лопиталя:
Эту неопределенность можно раскрыть по правилу Лопиталя: