
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Краткие теоретические сведения по основнымСтр 1 из 9Следующая ⇒
КРАТКИЕ ТЕОРЕТИЧЕСКИЕ СВЕДЕНИЯ ПО ОСНОВНЫМ ТЕМАМ ДИСЦИПЛИНЫ
Предмет и задачи микробиологии
Микробиология (от греч. micros – малый, bios – жизнь, logos – учение) изучает строение, жизнедеятельность, закономерности и условия развития организмов малых размеров, видимых с помощью оптических микроскопов. Эти организмы названы микроорганизмами или микробами. К ним относят: бактерии, микроскопические грибы, дрожжи, водоросли, простейшие, вирусы, рикетсии (облигатные внутриклеточные паразиты). В 1886 году немецкий ученый Э. Геккель предложил для царства микробов (третьего царства живых существ) название протисты от греческого слова protistos – самый первый. Протисты подразделяются на высших или эукариотов (гр. karion – ядро), клетки которых сходны с растительными и животными клетками, и низших – прокариотов (бактерии и сине-зеленые водоросли). Вирусы, как неклеточные частицы противопоставляются всем остальным организмам, так как не способны к самостоятельному размножению, а только внутри живых клеток. Микроорганизмы широко распространены во всех областях биосферы (почве, воде, воздухе). Множество живет как на поверхности, так и внутри тела животных и человека, на растениях, на окружающих предметах, пищевых продуктах и т.д. Высокая выносливость и легкая приспособленность к различным условиям существования, способность к быстрому размножению делают их неотъемлемыми невидимыми спутниками, участниками самых различных процессов: круговорота веществ, в плодородии почв, почвообразовании, деструкции ксенобиотиков. С их жизнедеятельностью связано образование каменного угля, нефти, некоторых руд, торфа; в производстве самых различных веществ (ацетона, этилового спирта, органических кислот, витаминов и т.д.). Задачи современной микробиологии настолько разнообразны и специфичны, что из неё выделились ряд дисциплин – медицинская, ветеринарная, сельскохозяйственная, геологическая, нефтяная, космическая и промышленная микробиологии.
Морфология и химический состав клетки – основного Объекта биотехнологии
Морфология. Основными формами бактерий являются шаровидная (монококки – одиночные, диплококки – попарно,тетракокки – четыре, стрептококки – длинные цепочки, сарцина – из 8 клеток, расположенных в два яруса один над другим, стафилококки – напоминающие грозди винограда). Палочковидные – цилиндрические. Извитые и изогнутые бактерии различаются по длине, толщине и степени изогнутости (вибрионы, спириллы, спирохеты). Помимо истинных бактерий имеются нитчатые бактерии, слизистые – миксобактерии, микроскопические грибы – актиномицеты, риккетсии. Размеры и форма тела могут изменяться под влиянием условий среды. Размеры малы и измеряются в мкм (микрометрах),1мкм = 1*103 мм.
Изогнутые формы бактерий все подвижные. Кокковые не способны к передвижению. Среди палочковидных имеются подвижные и неподвижные формы. Передвижение осуществляется жгутиками. К характерным особенностям бактерий относится способность чрезвычайно быстро размножаться: через каждые 20-30 минут их количество может удваиваться. Спорообразование происходит исключительно у палочковидных бактерий. Спорообразование процесс сложный и малоизученный. Спорообразование – это процесс приспособления к переживанию неблагоприятных условий среды. Спора развивается из части протоплазмы (цитоплазма с ядерным материалом) материнской вегетативной клетки. Диаметр спор у некоторых бактерий превышает ширину клетки, вследствие чего форма спорообразующих клеток изменяется. Клетки приобретают форму веретена (клостридиум), если спора расположена в центре клетки, или в виде барабанной палочки (плектридиум), когда спора находится на конце клетки. Спорообразующие бактерии аэробные и факультативно-анаэробные называются бациллами, анаэробные - клостридиями. Спорообразующие бактерии могут переходить в аспорогенные формы. Химический состав микроорганизмов. Для понимания процессов обмена веществ необходимо знать химический состав их тела. Он сходен с химическим составом тела животных и растений. Важнейшие элементы, которые преобладают в составе клетки – это водород – H, азот – N, сера – S, фосфор – P, калий – K, магний – Mg, кальций – Ca, железо – Fe. Первые четыре элемента называются органогенными, так как составляют основу органического вещества. На них приходится 90-97% сухого вещества (таблица 1).
Таблица 1 Среднее количество отдельных элементов в клетках микроорганизмов
Остальные элементы называются зольными или минеральными (3-10%). Микроэлементы: медь – Cu, цинк – Zn, марганец – Mn, молибден – Mo и другие. Они выполняют важную роль в процессе жизнедеятельности. Вода занимает в микробной клетке 75-90% их массы. Часть ее находится в связанном состоянии (с белками, углеводами и другими) и входит в состав клеточной структуры. Остальная вода находится в свободном состоянии и участвует во многих химических реакциях. Органические вещества. Сухое вещество тела микроорганизма не превышает 15-20%. На органические соединения (белки, жиры, углеводы) приходится до 85-95%. Пигменты (красящие вещества) содержаться главным образом в клеточном соке и обуславливают окраску бактерий, дрожжей, грибов. У некоторых бактерий они выполняют роль хлорофилла зеленых растений. Некоторые пигменты играют важную роль в процессе дыхания. Минеральные вещества встречаются в клетках микроорганизмов в виде хлоридов, сульфатов, фосфатов и играют большую роль в регуляции осмотического давления и коллоидного состава цитоплазмы, влияют на скорость и направление биохимических реакции, протекающих в клетке, являются стимуляторами роста и активаторами ферментов.
Ферменты микроорганизмов Все разнообразие биохимических процессов, протекающих в животном организме, совершается при участии биологических катализаторов – ферментов или энзимов. Ферменты открыты в 1814 году К.С Кирхгофом. Ферменты – это высокомолекулярные белки, образующие в воде коллоидные (гр. kolla – клей) растворы. Их можно разделить на 2 группы. 1. Протеины. Это однокомпонентные ферменты, состоящие только из белка. Роль активной группы, ответственной за каталитические свойства, могут выполнять SH-группы и остатки отдельных аминокислот. 2. Протеиды. Это двухкомпонентные ферменты, состоящие из белка и небелковой простетической группы. В простетическую группу входят витамины или их производные, различные металлы, азотистые основания. Простетическая группа обуславливает каталитическую способность фермента, а белковая – специфичные свойства фермента. Связь у простетической группы с белком не одинакова и имеет различную прочность. У некоторых ферментов простетическая группа легко отделяется от белков и может вступать во временную связь с другими белками. Такая простетическая группа называется ко-ферментом. Особенностью ферментов является их высокая активность и субстратная специфичность. По образному выражению Э. Фишера: «Субстрат и фермент подходят к друг другу как ключ к замку». Ферменты отличаются неустойчивостью к воздействиям факторов внешней среды: температура, лучистая энергия, рН среды, химические вещества, концентрация субстрата. Ферменты теряют свою активность (инактивируются), что во многом объясняется их белковой природой. Ферменты в клетке локализованы в различных ее структурах: митохондриях, мезосомах, цитоплазматической мембране. Некоторые ферменты выделяются в среду – экзоферменты, способствуя внеклеточному перевариванию пищи, подготавливая ее тем самым к поступлению в клетку. Эндоферменты не выделяются в окружающую среду. Они адсорбированы клеточными структурами и участвуют во внутриклеточных процессах обмена веществ.
В 1966 году на «Международном биохимическом союзе» специальной комиссией разработана классификация ферментов, согласно которой ферменты делятся на 6 классов по типу катализируемой ими реакции. 1. Оксидоредуктазы. Они катализируют окислительно-восстановительные реакции процессов дыхания, брожения. Например, дегидрогеназы, пероксидазы, каталазы. 2. Трансферазы – ферменты переноса атомных группировок от одного соединения к другому. Например, фосфотрансферазы, аминотрансферазы. 3. Гидролазы. Они расщепляют сложные органические вещества по типу гидролитических реакций с присоединением воды. Например, полиазы, олигазы, целлюлазы. 4. Лиазы. Они катализируют реакции негидролитического расщепления, которые сопровождаются отщеплением тех или иных групп. Например, декарбоксилазы. 5. Изомеразы. Катализируют внутримолекулярное перемещение радикалов, атомов, атомных группировок различных органических соединений. 6. Лигазы (синтетазы). Катализируют реакции соединения двух молекул, которые обычно протекают с расщеплением пирофосфатных связей. Например, карбоксилазы.
Типы питания
Углеродное питание. В зависимости от того, что является источником углерода микроорганизмы подразделяются на автотрофов (фототрофы и хемотрофы) и гетеротрофов. Фототрофы (фотосинтетики) – организмы, для которых источником энергии служит свет, источником углерода - углекислый газ, источник электронов – вода. Это, например, водоросли, зеленые и пурпурные серобактерии. Хемотрофы (хемосинтетики) получают энергию за счет окислительно-восстановительных реакций, а источником углерода является углекислый газ. Гетеротрофы – в качестве источника углерода используют органические вещества; источником энергии – окислительно-восстановительные реакции; источником электронов – органические вещества. Большинство живет за счет органических веществ животного и растительного происхождения (бактерии, грибы, дрожжи). Сапрофиты (гр. sapros – гнилой) разлагают органические вещества. Некоторые гетеротрофы – паразиты (паратрофы) – развиваются только в теле других организмов, питаясь органическим веществами, входящими в состав белков клетки хозяина. Гетеротрофы наряду с органическими соединениями используют и углекислый газ, вовлеккая его в обмен веществ. Углекислый газ служит дополнительным источником углерода для биосинтеза веществ тела.
Специфичность отношений микроорганизмов к источнику углеродной пищи определяется круговоротом углерода в природе. Азотное питание. Азот необходим для синтеза белка, нуклеиновых кислот и других азотистых веществ. Источники азота у микробов разнообразны. Паразиты используют азот хозяина. Сапрофиты не могут синтезировать белки из простых соединений, а только используя сложные органические вещества (пептоны, пептиды, полный набор аминокислот). Некоторым сапрофитам нужны только несколько аминокислот. Используемые аминокислоты дезаминируются, т.е. распадаются с потерей аммиака. Дезаминирование бывает: - восстановительное R – СН(NH2) – СООН + 2H → R – CH2 – COOH + NH3 - окислительное R – СН(NH2) – СООН + ½ О2 → R – CO – COOH + NH3 - гидролитическое R – СН(NH2) – СООН + H2O → R – CH(OH) – COOH + NH3 Синтез новых аминокислот может происходить путем переаминирования – перенос аминогрупп с аминокислоты на кетокислоту при участии фермента аминотрансферазы. R – СН(NH2) – СООН + СН3 – СО – СООН → R– СО – СООН + СН3 – СН(NH2) – СООН Многие микроорганизмы в качестве источника азота используют нитраты NO3-, реже нитриты NO2-. Есть бактерии и грибы, которые способны использовать свободный азот атмосферы. Усвоение зольных элементов. Зольные элементы: сера, фосфор, калий, кальций, магний, железо. Хотя потребность в них незначительна, но недостаток их может привести к гибели организма. Микроорганизмы усваивают их из минеральных солей.
Извлечения энергии клеткой
Клетка способна сохранять весьма специфичную структуру только благодаря непрерывному потреблению энергии, иначе она теряет структуру и переходит в неупорядоченное и лишенное организации состояние. Установлено, в каком органе клетки происходит тот или иной биохимический процесс извлечения энергии. Фотосинтез обеспечивают хлоропласты, которые содержат зеленый пигмент хлорофилл. Дыхание обеспечивают митахондрии и мезосомы. Специфические молекулы в клетке переносят энергию, полученную в процессах фотосинтеза и дыхания, от центров фотосинтеза и дыхания во все участки клетки, где протекают процессы, связанные с потреблением энергии. Этими молекулами является АТФ. АТФ → АДФ + неорганический фосфат. В процессе этой реакции свободная энергия молекул АТФ превращается в тепловую. Значительная часть энергии молекулы АТФ сохраняется благодаря фосфорилированию молекулы-акцептора, которая приобретает возможность участвовать в процессах, протекающих с потреблением энергии. АДФ – это разряженная форма АТФ, бедная энергией. Первичная зарядка АТФ происходит при фотосинтезе, вторичная – в процессе дыхания.
Фотосинтез
При фотосинтезе энергия солнечного превращается в химическую и расходуется на «зарядку» АДФ, на образование АТФ. В клетках содержится зеленый пигмент – хлорофилл. Солнечный свет с определенными длинами волн поглощается хлорофиллом. Световой цикл фотосинтеза – цикл Д.Арноноа.
Рис. 1 Схема превращения энергии света.
Квант света (рис.1), попадая в молекулу хлорофилла, поглощается. Под влиянием поглощенного света хлорофилл приходит в триплетное (возбужденное) состояние. Поглощенная энергия переводит электроны в сложной молекуле хлорофилла с основного энергетического уровня на более высокий. Богатые энергией электроны не возвращаются в свое нормальное состояние. Вместо этого они отрываются от молекулы хлорофилла и переносятся специфическими молекулами-переносчиками электронов (А, В, С, Д, Е), которые передают электроны друг другу по замкнутой цепи. Проделывая путь вне молекулы хлорофилла, возбужденные электроны передают поглощенную энергию от одного переносчика другому и возвращаются на свой основной уровень в молекуле хлорофилла. После этой процедуры молекула хлорофилла оказывается вновь готовой к поглощению второго кванта света. Энергия, отданная электрону, используется на образование АТФ из АДФ, т.е. на зарядку АТФ от системы фотосинтеза клетки. Этот процесс называется окислительным фосфорилированием. Помимо связывания солнечной энергии в процессе фотосинтеза происходит еще и фотолиз воды (рис.2).
 


Рис.2 Схема фотолиза воды.
В настоящее время полагают, что некоторые из «горячих» электронов вместе с ионами водорода происходят из молекулы воды. Ион водорода вызывает восстановление одного из переносчиков электронов – трифосфопиридиннуклеотид (ТПН). В восстановленной форме – ТПН-Н. Одним из побочных продуктов восстановления ТПН служит гидроксил ион ОН-. Этот ион отдает свой электрон одному из цитохромов в цепи фотохимических реакции, и конечным продуктом оказывается молекулярный кислород. Синтез глюкозы. Для течения этой реакции необходимы два соединения – АТФ и ТПН-Н, образующиеся только на свету и служащие источником энергии (АТФ) и восстанавливающим агентом (ТПН-Н). Главным промежуточным продуктом в сложном цикле реакций служит рибулозодифосфат (РДФ), который присоединяет к себе углекислый газ и образуется две молекулы 3-фосфоглицерата. Фосфоглицерат восстанавливается при участии ТПН-Н и в результате ряда реакций превращается в глюкозу. В результате ряда реакций – пентозный шунт – вновь образуется РДФ (рис.3). Таким образом, молекула глюкозы представляет конечный продукт фотосинтеза.
Рис.3 Схема синтеза глюкозы.
Дыхание.
Дыхание – окисление органического вещества. Аэробные микроорганизмы (грибы, водоросли, многие бактерии, дрожжи) используют чаще всего в качестве энергетического материала углеводы (главным образом, глюкозу). Многие аэробы окисляют органические вещества полностью, выделяя в виде конечных продуктов углекислый газ и воду. При полном окислении глюкозы освобождается вся потенциальная энергия, которая была аккумулирована в молекуле сахара при фотосинтезе. Процесс окисления глюкозы в клетке делится на две основные фазы: гликолиз (анаэробный процесс) и цикл Кребса (аэробный процесс). Гликолиз – первая или подготовительная фаза. В результате этого процесса происходит расщепление глюкозы до молочной кислоты. Процесс состоит из 11 ступеней, каждая из которых катализируется своим ферментом. 1. Фосфорилирование молекулы глюкозы глюкоза + АТФ 2. Изомеризация глюкоза-6-фосфат 3. Фосфорилирование молекулы фруктозы фруктоза-6-фосфат+АДФ 4. Расщепление фруктозы-1,6-дифосфат фруктоза-1,6-дифосфат альдегид 5. Изомеризация фосфатдиоксиацетон 6. Присоединение фосфорной кислоты и 2ТПН 3фосфоглицериновый + 2Н3РО4 альдегид 7. Подзарядка АДФ 1,3-дифосфоглицерат+ АДФ+2Р кислота 8. Отнятие фосфора 3фосфоглицериновая кислота 9. Отнятие воды 2фосфоглицериновая кислота 10. Подзарядка АДФ 2энолфосфопируват + 2АДФ пировиноградная кислота (кетоформа) 11. Восстановление кетоформы в энольную форму пировиноградной кислоты 2 СН3 – СО - СООН молочная кислота Эти начальные этапы окисления глюкозы до пировиноградной кислоты проходят однотипно как у аэробов, так и у анаэробов без участия кислорода. После расщепления глюкозы до пировиноградной кислоты аэробные клетки продолжают извлекать большую часть оставшейся энергии в процессе дыхания. Пировиноградная кислота претерпевает ещё более сложный ряд превращений. Цикл превращений пировиноградной кислоты был описан Г. Кребсом в 1937 году. Цикл Кребса.
Цикл Кребса важен не только для процессов дыхания (получение энергии), но и для биосинтеза. Он снабжает клетку веществами, легко превращающихся в аминокислоты, белки, жиры. При дыхании идут два процесса: перенос электронов водорода с одного соединения на другое (дыхательная цепь → кислород); перенос фосфорной группы с АДФ на АТФ.
Брожение
Анаэробные микроорганизмы получают необходимую энергию в процессе брожения. К брожению в строгом смысле слова относят процессы получения энергии, при которых отщепленный от субстрата водород переносится на органический субстрат, являющийся терминальным акцептором. Кислород в процессе брожения не участвует. Брожение следует отличать от анаэробного дыхания. В зависимости от конечного акцептора водорода различают: дыхание – конечным акцептором является кислород; брожение – конечным акцептором – органические вещества (спирты, масляная, молочная кислоты); анаэробное дыхание – процесс протекает без участия кислорода, водород передается через дыхательную цепь терминальному акцептору – NO2-, SO4-2. В зависимости от характерных или преобладающих продуктов, выделяющихся в процессе брожения, различают: спиртовое (рис.4), молочнокислое, маслянокислое, пропионовое, уксусное, а также брожение, приводящее к образованию метана. Энергетическим материалом при брожении чаще всего служат углеводы.
2 НАД+ 2 НАД-Н (+Н+)
Рис.4. Превращение пировиноградной кислоты в этанол при сбраживании глюкозы.
Физиология роста. Под ростом понимают увеличение количества живого вещества, сопровождающегося увеличением и делением клеток. У многоклеточных микроорганизмов увеличиваются размеры организма, у одноклеточных - число клеток и масса. Концентрация клеток – определенное количество клеток в гомогенной (однородной) суспензии (число клеток в 1 мл). Плотность – отношение массы клеток к объему (мг/дм3). Константа скорости деления – число клеточных делений за 1 час. Время генерации – время, за которое число клеток в бактериальной популяции удваивается. Рост бактерии в статистической культуре. Кривая роста. При внесении бактерий в питательную среду они растут до тех пор, пока содержание каких-либо необходимых компонентов не достигнет минимума, после этого рост прекращается. Если в среду не добавлять питательные вещества и не удалять метаболиты, то получим статистическую бактериальную культуру, т.е. популяцию в ограниченном жизненном пространстве. Отложив на оси ординат число клеток в экспоненциально растущей популяции, а на оси абсцисс – время, то получим кривую роста (рис.5), которая описывается экспоненциальным законом.
 
 I II III IV I II III IV
время t
Кривая позволяет различить несколько фаз роста, сменяющих друг друга в определенной последовательности. I. Лаг-фаза – начальная фаза роста. Она охватывает промежуток времени между инакуляцией (заражением) и достижением максимальной скорости роста. Продолжительность зависит от возраста инакулянта, пригодности среды, адаптации к новым учловиям. II. Экспоненциальная фаза (логарифмическая) характеризуется постоянной максимальной скоростью деления клеток. Зависит от вида бактерий. III. Стационарная фаза наступает тогда, когда число клеток перестает увеличиваться. Переход к этой фазе происходит постепенно. Скорость роста ограничивается не только количеством субстрата, но и большой плотностью бактериальной популяции, низким парциальным давлением, токсичностью продуктов обмена. В стационарной фазе может происходить использование запасных питательных веществ, распад клеточных субъединиц, синтез ферментов. IV. Фаза отмирания. Отмирание клеток и причины их гибели в нормальных питательных средах изучены недостаточно.
12. Инженерные основы биотехнологии.
Биотехнология – процесс производства, протекающий с использованием живых микроорганизмов. Биотехнология теснейшим образом связана с появлением генной инженерии. Биотехнологические процессы используются в производстве ценных биологически активных веществ (антибиотиков, гармонов и т.д.); для предотвращения загрязнения окружающей среды (биологическая очистка); защиты растений от вредителей и болезней, в производстве белков и аминокислот, лекарственных препаратов (интерферона, инсулина, гормонов роста). Биотехнологическими объектами являются микроорганизмы, клеточные ферменты, антитела. Цель – применение генной инженерии на практике. Генная инженерия – отрасль молекулярной биологии, изучающая направленные изменения биологической информации клетки для получения живых существ с запрограммированными фенотипическими признаками. Фенотипические признаки – совокупность всех свойств и признаков особи на определенной стадии развития, сформировавшиеся в результате взаимодействия генотипа с окружающей средой. Генотип – совокупность всех генов, присутствующих в данной особи. Синтез генов – выделение гена из клетки-донора; присоединение гена к ДНК (вектору), способная вести его в клетке гена в геном реципиента. Геном – совокупность хромосом, содержащие единицы наследственной информации – гены. Наследственные или целенаправленные изменения в генетическом аппарате называются мутациями. В биотехнологии используются различные аппараты (биологические реакторы). 1. Идеальный проточный реактор с полным перемешиванием (ПРПП). Предназначены для изучения роста культур клеток, поэтому их часто называют хемостатами. Перемешивание в таких реакторах осуществляется с помощью мешалки. Полное перемешивание обеспечивает однородность состава среды и одинаковую концентрацию кислорода. Такие биореакторы обеспечивают достаточно однородные условия, что необходимо для изучения кинетики соответствующих процессов. Кинетика изучает механизмы скорости микробиологических процессов, основы управляемого количественного биосинтеза. Математически описывает протекающие биопроцессы во времени. ПРПП используются для изучения и определения параметров реакторов с неполным перемешиванием. 2. В крупномасштабном производстве необходимыми и целесообразными являются периодические процессы, отличающиеся непостоянным несбалансированным ростом клеток, различными метаболитическими процессами, и их активностью в разные периоды. Для таких процессов используются реакторы периодического действия. 3. Идеальный трубчатый реактор полного вытеснения (ТРПВ). В трубчатом реакторе стационарное состояние сохраняется в элементарном объеме потока, перпендикулярном оси реактора. Система сигрегирована (лат. segregare – отделять, удалять), и каждый элемент системы ведет себя как реактор периодического действия. ТРПВ обеспечивает высокую степень превращения субстрата на выходе из реактора, по сравнению с ПРПП. В тоже время в такой реактор необходимо вносить посевной материал. 4. Реакторы с неподвижным слоем биокатализатора. Работа такого реактора определяется свойствами, применяющихся биокатализаторов. В основном применяются катализаторы на основе иммобилизованных (лат. immobilis – неподвижный) клеток. Иммобилизованные клетки обладают рядом преимуществ: создают возможность для осуществления непрерывного процесса (окисления вещества) без вымывания микроорганизмов и изменения генетической природы; позволяют непрерывно отделять продукты и удалять ингибирующие вещества. Примение иммобилизованных клеток в биореакторах дает возможность расширить временной диапазон, в течение которого клетки проявляют свои каталитические свойства. 5. Биореакторы с псевдоожиженным слоем катализатора. Процессы осуществляются в реакторах колонного типа. Псевдоожиженный слой предполагает наличие трех фаз: газ, жидкость, твердое тело. Поток жидкости в таком реакторе направлен снизу вверх по высокому вертикальному цилиндру. Частицы нерастворимого биокатализатора (скопление микроорганизмов, частицы иммобилизованных клеток, ферментов) суспендируются, увлекаемые восходящим потоком жидкости. Частицы катализатора в верхней расширенной части реактора прекращают подъем и вновь поступают в башню. Такие реакторы используются в непрерывных процессах пивоварения. 6. Реакторы с неподвижным слоем катализатора и со струйным течением жидкости. Область применения таких реакторов – обработка сточных вод с помощью биологических капельных фильтров, производство винного уксуса, в нефтеперерабатывающей промышленности для гидрокрекинга и гидроочистки.
I Микробиология Тема 1 Светопольная микроскопия Светопольная микроскопия
Все световые биологические микроскопы отечественного производства можно разделить на три группы: биологические упрощенные, биологические рабочие, биологические исследовательские. Наиболее распространенной моделью является микроскоп МБР-1.
Устройство микроскопа
Микроскоп (рис. 1) состоит из механической и оптической частей. Механическая часть микроскопа включает подковообразный башмак 9, коробку с микромеханизмом 10, предметный столик 11, револьвер на салазках 12, кронштейн конденсатора 13. Коробка микромеханизма несет, с одной стороны, направляющую для тубусодержателя. Внутри коробки находится механизм фокусировки микроскопа. На боковых поверхностях коробки расположены рукоятки микромеханизма 1, служащего для точной фокусировки объектива. Слева на оси рукояток закреплен барабан со шкалой, разделенной на 50 частей. Цена одного деления барабана 0,002 мм. Один оборот барабана соответствует перемещению тубуса на 0,1 мм. Микромеханизм перемещает тубус вместе с механизмом грубой и точной фокусировки, по часовой стрелке тубус микроскопа опускается, при вращении против часовой стрелки – поднимается. Предметный столик 2 укреплен на специальном кронштейне, закрепленном на верхней части коробки микромеханизма. Верхняя часть предметного столика может вращаться, если опустить стопорный винт 3. Столик вращают рукой за накатанную торцовую часть. С помощью винтов 2, находящихся справа и слева на корпусе кронштейна, столик перемещается (центрируется) на 8 мм, что позволяет подвести нужный участок препарата в поле зрения. На поверхности столика расположено семь отверстий. Четыре крайних отверстия служат для установки пружинных клемм 8, прижимающих препарат, а три средних отверстия – для крепления накладного препаратоводителя. Дугообразный тубусодержатель в нижней своей части имеет рукоятку макровинта 4, используемую для грубой фокусировки микроскопа. У верхней части тубусодержателя снизу находится головка для крепления револьвера, а сверху – специальное посадочное гнездо для крепления сменных тубусов: наклонного – для визуальных исследований и прямого – для фотографических работ. Револьвер 12 имеет гнезда с резьбой для крепления объективов. Вращением револьвера осуществляется смена объективов. Отверстия для объективов на револьвере отцентрированы относительно оси тубуса с такой точностью, что при переходе от слабого объектива к более сильному и при повороте револьвера по часовой стрелке точка препарата, установленная при слабом объективе в центре поля зрения окуляра 16, всегда остается в поле зрения более сильного объектива.
Рис. 1 Общий вид иммерсионного микроскопа. 1 – рукоятка микромеханизма; 2 – центрировочный винт для установки препарата; 3 – стопорный винт; 4 – рукоятка грубой фокусировки; 5 – винт для укрепления тубуса; 6 – рукоятка перемещения диафрагмы; 7 - стопорный винт конденсора; 8 – пружинные клеммы; 9 – подковообразный башмак; 10 – коробка с микромеханизмом; 11 – предметный столик; 12 – револьвер на салазках; 13 – кронштейн конденсора; 14 – конденсор; 15 – объектив; 16 – окуляр; 17 – головка тубосодержателя; 18 – зеркало; 19 – ирисовая апертурная диафрагма; 20 – призма; 21 – рукоятка перемещения конденсора; 22 – рукоятка ирисовой диафрагмы конденсора; 23 – револьвер. Кронштейн 13 имеет цилиндрическую гильзу для конденсора. Конденсор 14 крепится в гильзе винтом 7, расположенным сбоку кольца кронштейна. Под кронштейном конденсора в коробке микромеханизма укреплена вращающаяся вилка для зеркала.
Оптическая часть микроскопа состоит из осветительной и наблюдательной систем. Осветительная система находится под предметным столиком. К ней относится зеркало 18 и конденсор 14 с ирисовой апертурной диафрагмой 19. Зеркало служит для отражения идущих от источника световых лучей по направлению к объективу и через него – внутрь микроскопа. Одна сторона зеркала плоская, другая – вогнутая. При работе с конденсором следует пользоваться только плоским зеркалом независимо от источника света. Вогнутое зеркало может применяться при работе без конденсора с объективами малых увеличений.
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-02-08; просмотров: 339; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.17.6.75 (0.156 с.) |






 глюкоза-6-фосфат + АДФ
глюкоза-6-фосфат + АДФ фруктоза-6-фосфат
фруктоза-6-фосфат фруктоза-1,6-дифосфат+АДФ
фруктоза-1,6-дифосфат+АДФ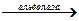 фосфатдиоксиацетон + фосфоглицериновый
фосфатдиоксиацетон + фосфоглицериновый 3 фосфоглицериновый альдегид
3 фосфоглицериновый альдегид 1,3-дифосфоглицерат + 2ТПН-Н
1,3-дифосфоглицерат + 2ТПН-Н 3фосфоглицериновая + 2АТФ
3фосфоглицериновая + 2АТФ 2фосфоглицериновая кислота
2фосфоглицериновая кислота 2энолфосфопируват + Н2О
2энолфосфопируват + Н2О 2 СН3 – СО - СООН + 2АТФ
2 СН3 – СО - СООН + 2АТФ 2 СН3 – СН(ОН) – СООН + 2ТПН
2 СН3 – СН(ОН) – СООН + 2ТПН По видимому,в процессе цикла Кребса электроны при участии ферментов отрываются от промежуточных продуктов и передаются ряду молекул-переносчиков, объединенных под общим названием «дыхательная цепь». Эта цепь ферментных молекул представляет конечный общий путь электронов. В последнем звене цепи электроны соединяются с кислородом и образуют воду.
По видимому,в процессе цикла Кребса электроны при участии ферментов отрываются от промежуточных продуктов и передаются ряду молекул-переносчиков, объединенных под общим названием «дыхательная цепь». Эта цепь ферментных молекул представляет конечный общий путь электронов. В последнем звене цепи электроны соединяются с кислородом и образуют воду.


 С6Н12О6 2 СН3 – СО – СООН
С6Н12О6 2 СН3 – СО – СООН СО2
СО2
 2 СН3 – СН2ОН СН3 – СОН
2 СН3 – СН2ОН СН3 – СОН lg N (число клеток)
lg N (число клеток)








