Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Центральная нервная система. . 32
6.1. Спинной мозг. 33 Рис. 20. Спинной мозг (вид с дорсальной стороны): 33 Рис. 21. Оболочки спинного мозга (шейный отдел): 34 Рис. 22. Оболочки спинного мозга на поперечном разрезе: 34 Рис. 23. Схема расположения сегментов спинного мозга: 35 Рис. 24. Схема поперечного среза спинного мозга: 36 Белое вещество. 37 Восходящие пути. 37 К нисходящим путям относятся: 37 6.2. Головной мозг. 38 6.2.1. Продолговатый мозг. 38 Рис. 25. Ствол мозга с дорсальной стороны. Мозжечок убран: 39 Рис. 26. Ствол мозга с вентральной стороны (римскими цифрами обозначены места выхода черепно-мозговых нервов): 40 Рис. 27. Ствол мозга сбоку. Мозжечок убран (римскими цифрами обозначены места выхода черепно-мозговых нервов): 41 Языкоглоточный нерв. 41 Блуждающий нерв. 41 Добавочный и подъязычный нервы.. 41 Рис. 28. Поперечный разрез продолговатого мозга. 42 Филогенез. 43 Онтогенез. 43 6.2.2. Задний мозг. 43 6.2.2.1. Варолиев мост. 43 В филогенезе. 44 В онтогенезе. 44 6.2.2.2. Мозжечок. 45 Рис. 29. Расположение мозжечка на стволе мозга (часть ткани мозжечка убрана): 45 Рис. 30. Мозжечок: 46 Рис. 31. Ядра мозжечка: 47 Рис. 32. Схема строения коры мозжечка: 47 Филогенез. 48 Онтогенез. 48 6.2.3. Средний мозг. 48 Рис. 33. Сагиттальный срез ствола мозга: 49 Рис. 34. Разрез среднего мозга (изображены примыкающие к нему части промежуточного мозга): 49 Рис. 35. Поперечный разрез через ножки мозга: 50 Филогенез. 50 Онтогенез. 51 6.2.4. Промежуточный мозг. 51 Рис. 36. Промежуточный и средний мозг (вид сверху): 51 6.2.4.1. Таламус. 51 Рис. 37. Схема основных ядерных групп промежуточного мозга: 52 6.2.4.2. Гипоталамус. 53 Рис. 38. Схема основных ядерных групп промежуточного мозга и среднего мозга: 53 6.2.4.3. Субталамус. 54 6.2.4.4. Эпиталамус. 54 6.2.4.5. Гипофиз 54 Нейрогипофиз. 54 Аденогипофиз (передняя доля) 54 Филогенез. 55 Онтогенез. 55 6.2.5. Конечный мозг. 56 Рис. 39. Схема взаимоотношений оболочек головного мозга: 56 Рис. 40. Система желудочков и цистерн мозга: 57 6.2.5.1. Базальные ганглии. 57 Рис. 41. Полушария мозга на разных уровнях горизонтального разреза: 58 Рис. 42. Топография базальных ядер конечного мозга и стволовых структур (вид слева спереди): 59 Рис. 43. Топография базальных ядер конечного мозга и стволовых структур (вид слева сзади): 59 6.2.5.2. Проводящие пути больших полушарий. 60 Проекционные пути. 60
Свод (fornix) 60 Рис. 44. Гиппокамп и свод. 61 Ассоциативными путями. 61 Короткие. 61 Длинные. 61 Комиссуральные пути. 61 Рис. 45. Проводящие пути больших полушарий (часть латеральной поверхности левого полушария удалена): 62 Рис. 46. Передняя комиссура и передняя часть височной доли (поперечный срез, вид сзади): 63 6.2.5.3. Кора. 63 Рис. 47. Фронтальный срез нижнего отдела височной доли левого полушария: 64 Древняя и старая кора. 64 Гиппокамп (hippocampus), или аммонов рог, 64 Лимбической системы мозга (Латинское слово limbus означает кайма, край.) 65 Рис. 48. Схема строения лимбической системы: 65 Рис. 49. Схема цитоархитектонического строения новой коры головного мозга: 66 Новая кора. 66 ♦ I слой — самый поверхностный, толщиной около 0,2 мм, называется молекулярным.. 66 ♦ II слой — наружный зернистый (lamina granulans externa). 66 Рис. 50. Пирамидный нейрон коры: 67 ♦ III слой — пирамидный (lamina pyramidalis), 67 ♦ IV слой — внутренний зернистый (lamina granulans internus). 67 ♦ V слой — ганглионарный (lamina ganglionaris) 67 ♦ VI слой коры — полиморфный (lamina multiformis). 67 Рис. 51. Латеральная поверхность левого полушария (Сильвиева борозда раздвинута): 68 Филогенез. 69 Онтогенез. 70 ОРГАНЫ ЧУВСТВ.. 70 7.1. Зрительная система. 71 Рис. 53. Схема горизонтального сечения правого глаза: 71 Глазное яблоко. 71 Роговица (cornea)— это выпуклая пластинка блюдцеобразной формы.. 71 Хрусталик. 72 Рис. 54. Структура сетчатки: 72 Рис. 55. Препарат вентральной поверхности мозга (височные доли и ствол сняты, средний мозг рассечен): 74 Рис. 56. Схема проводящих путей зрительного анализатора: 75 7.2. Слух и равновесие. 76 7.2.1. Органы слуха. 76 Наружное ухо. 76 Рис. 57. Схема строения среднего и внутреннего уха: 76 Среднее ухо. 76 Внутреннее ухо. 76 Костный лабиринт. 77 Костная улитка. 77 Рис. 58. Схема строения Кортиевого органа: 77 Рис. 59. Волосковая клетка: 78 Филогенез. 78 Онтогенез. 78 7.2.2. Вестибулярная система. 78 Отолитовый аппарат. 79 Рис. 60. Схема строения макулы отолитового аппарата: 79 7.3. Вкусовая система. 80 Рис. 61. Схема вкусовой почки, погруженной в сосочек языка: 80 Рис. 62. Схема локализации чувствительных структур на поверхности языка человека: 81 Филогенез. 81
Онтогенез. 81 7.4. Обонятельная система. 81 Рис. 63. Строение обонятельного эпителия: 82 Онтогенез. 83 7.5. Кожная рецепция. 83 Рис. 64. Кожные рецепторы: 83 Рис. 65. Соединение путей кожных и висцеральных рецепторов в спинном мозге: 84 Болевая чувствительность (ноцицепция) 84 7.6. Проприоцепция и интероцепция. 84 Проприоцептивный анализатор. 84 Интероцептивный анализатор. 85 Приложение. Словарь латинских терминов. 86 Обозначения направлений. 86 Общие термины.. 86 Список литературы.. 88 Содержание. 89
Иллюстрации Рис. 1. Формирование нервной ткани: 11 Рис. 2. Нейрон: 12 Рис. 3. Схема миелинизации аксонов: 13 Рис. 4. Строение синапса: 13 Рис. 5. Типы нейронов: 16 Рис. 6. Схема взаиморасположения элементов нервной ткани: 17 Рис. 7. Поперечный срез через нерв (а) и нервный ствол (б): 17 Рис. 8. Схема строения диффузной нервной системы кишечнополостного животного: 19 Рис. 9. Схема строения диффузностволовой нервной системы турбеллярии: 19 Рис. 10. Схема строения ортогональной нервной системы ресничного червя (передний конец): 20 Рис. 11. Схема строения узловой нервной системы насекомого: 21 Рис. 12. Схема строения головного мозга насекомого (пчела). Левая половина — его сечение: 22 Рис. 13. Схема строения ганглионарной нервной системы пластинчатожаберного моллюска (беззубка): 22 Рис. 14. Схема строения нервной системы головоногого моллюска (осьминог): 23 Рис. 15. Схема развития плаща конечного мозга (обозначен черным) в сравнении с остальными структурами мозга в ряду позвоночных: 24 Рис. 16. Закладка нервной трубки (схематичное изображение и вид на поперечном срезе): 26 Рис. 17. Пренатальное развитие нервной системы человека: 26 Рис. 18. Этапы развития головного мозга человека. 28 Рис. 19. Схема строения вегетативной нервной системы. Римскими цифрами обозначены соответствующие пары черепно-мозговых нервов. 30 Рис. 20. Спинной мозг (вид с дорсальной стороны): 33 Рис. 21. Оболочки спинного мозга (шейный отдел): 34 Рис. 22. Оболочки спинного мозга на поперечном разрезе: 34 Рис. 23. Схема расположения сегментов спинного мозга: 35 Рис. 24. Схема поперечного среза спинного мозга: 36 Рис. 25. Ствол мозга с дорсальной стороны. Мозжечок убран: 39 Рис. 26. Ствол мозга с вентральной стороны (римскими цифрами обозначены места выхода черепно-мозговых нервов): 40 Рис. 27. Ствол мозга сбоку. Мозжечок убран (римскими цифрами обозначены места выхода черепно-мозговых нервов): 41 Рис. 28. Поперечный разрез продолговатого мозга. 42 Рис. 29. Расположение мозжечка на стволе мозга (часть ткани мозжечка убрана): 45 Рис. 30. Мозжечок: 46 Рис. 31. Ядра мозжечка: 47 Рис. 32. Схема строения коры мозжечка: 47 Рис. 33. Сагиттальный срез ствола мозга: 49 Рис. 34. Разрез среднего мозга (изображены примыкающие к нему части промежуточного мозга): 49 Рис. 35. Поперечный разрез через ножки мозга: 50 Рис. 36. Промежуточный и средний мозг (вид сверху): 51 Рис. 37. Схема основных ядерных групп промежуточного мозга: 52 Рис. 38. Схема основных ядерных групп промежуточного мозга и среднего мозга: 53 Рис. 39. Схема взаимоотношений оболочек головного мозга: 56 Рис. 40. Система желудочков и цистерн мозга: 57 Рис. 41. Полушария мозга на разных уровнях горизонтального разреза: 58 Рис. 42. Топография базальных ядер конечного мозга и стволовых структур (вид слева спереди): 59
Рис. 43. Топография базальных ядер конечного мозга и стволовых структур (вид слева сзади): 59 Рис. 44. Гиппокамп и свод. 61 Рис. 45. Проводящие пути больших полушарий (часть латеральной поверхности левого полушария удалена): 62 Рис. 46. Передняя комиссура и передняя часть височной доли (поперечный срез, вид сзади): 63 Рис. 47. Фронтальный срез нижнего отдела височной доли левого полушария: 64 Рис. 48. Схема строения лимбической системы: 65 Рис. 49. Схема цитоархитектонического строения новой коры головного мозга: 66 Рис. 50. Пирамидный нейрон коры: 67 Рис. 51. Латеральная поверхность левого полушария (Сильвиева борозда раздвинута): 68 Рис. 53. Схема горизонтального сечения правого глаза: 71 Рис. 54. Структура сетчатки: 72 Рис. 55. Препарат вентральной поверхности мозга (височные доли и ствол сняты, средний мозг рассечен): 74 Рис. 56. Схема проводящих путей зрительного анализатора: 75 Рис. 57. Схема строения среднего и внутреннего уха: 76 Рис. 58. Схема строения Кортиевого органа: 77 Рис. 59. Волосковая клетка: 78 Рис. 60. Схема строения макулы отолитового аппарата: 79 Рис. 61. Схема вкусовой почки, погруженной в сосочек языка: 80 Рис. 62. Схема локализации чувствительных структур на поверхности языка человека: 81 Рис. 63. Строение обонятельного эпителия: 82 Рис. 64. Кожные рецепторы: 83 Рис. 65. Соединение путей кожных и висцеральных рецепторов в спинном мозге: 84
Светлой памяти нашего Учителя Григория Аствацатуровича Кураева посвящается
Предисловие Введение в учебные планы подготовки студентов-психологов курса анатомии центральной нервной системы (ЦНС) отражает очевидную необходимость в подобных знаниях. Особенность данного курса, по мнению авторов пособия — сочетание морфологии и отдельных аспектов онто- и филогенеза нервной системы, а также его логическая связь с последующими курсами: физиологии нервной системы, физиологии высшей нервной деятельности и т.д. Изложение курса анатомии центральной нервной системы студентам-психологам требует специфического подбора материала. С одной стороны, строение структур ЦНС необходимо описывать достаточно подробно, с другой — материал не должен быть перегружен множеством деталей анатомии мозга и латинской терминологией, что характерно для фундаментальных медицинских атласов и учебников по анатомии. Авторы попытались сохранить баланс между академичностью изложения курса и его доступностью.
Мы старались достаточно полно проиллюстрировать пособие, с тем чтобы по возможности облегчить понимание такого сложного материала, как строение центральной нервной системы. Кроме того, прилагается краткий словарь латинских терминов, сгруппированных согласно расположению отделов ЦНС. Внутри каждого раздела термины расположены исходя из соотношения между обозначаемыми анатомическими структурами. Знание латинских терминов поможет студентам разобраться в терминологии фундаментальных работ по анатомии. ОБЩИЕ СВЕДЕНИЯ Нервная система состоит из специфической возбудимой ткани — нервной ткани — и представлена центральным и периферическим отделами. Центральная нервная система находится под защитой костных образований скелета: черепной коробки, в которой расположен головной мозг, а также позвоночника, в спинномозговом канале которого расположен спинной мозг. К периферической нервной системе относятся нервы и нервные узлы. Выделяют соматическую и вегетативную части периферической нервной системы. Часть нервной системы, которая регулирует работу скелетных мышц, называют соматической (греч. soma — тело). Посредством соматической нервной системы человек может управлять движениями, произвольно вызывать или прекращать их. Часть нервной системы, регулирующую деятельность внутренних органов (сердца, желудка, желез и т.д.), называют вегетативной. Работа вегетативной нервной системы не подчиняется воле человека. Для обозначения взаимного расположения структур нервной системы анатомы пользуются специфическими терминами. Названия направлений основаны на их латинских названиях. Плоскость, проходящая вдоль по середине тела и делящая его на правую и левую половины, называется сагиттальной (sagittalis).Структуры, расположенные на спинной стороне тела, называются дорсальными (dors a lis — спинной), расположенные на брюшной стороне — вентральными (ventr a lis — брюшной). Структуры, лежащие по центру тела (близко к сагиттальной плоскости), называются медиальными (medi a lis — срединный), лежащие вбок от сагиттальной плоскости — латеральными (later a lis — боковой). Самые верхние точки структур называются апикальными (apic a lis — верхушечный), лежащие в основании — базальными (bas a lis).Направление к нижней части тела называется каудальное (caud a lis — хвостовой), а к головной части — ростральное (rostrum — клюв). НЕРВНАЯ ТКАНЬ Закладка нервной системы (рис. 1) начинается с формирования нервной пластинки, которая представляет собой полоску утолщенной эмбриональной эктодермы, расположенную над закладкой хорды. Нервная пластинка прогибается, ее края смыкаются, и образуется нервная трубка, которая отделяется от эктодермы и погружается под нее. В начале формирования стенка нервной трубки состоит из слоя цилиндрических клеток нейроэпителия, окружающего полость — центральный канал нервной трубки. По мере деления клеток стенка нервной трубки утолщается. Слой клеток, прилежащих к центральному каналу, называется эпендимным. Эти клетки дают начало почти всем клеткам нервной системы. Каждая зачатковая клетка делится на две дочерних. Одна из них мигрирует в вышележащие слои и становится нейробластом. Нейробласты претерпевают изменения, образуя характерные отростки, и дифференцируются в зрелые нервные клетки — нейроны.
Другие потомки зачатковых клеток остаются прикрепленными к внутренней мембране и образуют длинные радиальные отростки, достигающие наружной мембраны нервной трубки. Они называются спонгиобластами. Спонгиобласты играют значительную роль в формировании нервной ткани, так как именно по их отросткам мигрируют дифференцирующиеся нервные клетки. Ориентируясь на ход отростков спонгиобластов, нервные клетки формируют отростки и занимают свое окончательное местоположение, определяющее их будущие связи с другими нервными клетками и их функцию. Позже спонгиобласты дифференцируются в глиальные элементы. В дальнейшем часть спонгиобластов теряет связь с наружной мембраной нервной трубки: они остаются прикрепленными к внутренней мембране и образуют клеточную выстилку центрального канала и желудочков зрелого мозга — эпендиму. Эпендимные клетки имеют Рис. 1. Формирование нервной ткани: а — поперечный срез закладки головного мозга: 1 — полость желудочка; 2 — отросток радиальной глиальной клетки; 6 — увеличенный сегмент стенки нервной трубки: 3 — внутренняя поверхность; 4 — вентрикулярная зона (слой тел глиальных клеток); 5 — субвентрикулярная зона; 6 — промежуточная зона; 7 — корковая пластина; 8 — краевая зона; 9 — наружная поверхность; в — увеличенный фрагмент ткани, помеченный рамкой на б: 10 — ведущий отросток нейрона; 11 — мигрирующий нейрон; 12 — хвостовой отросток нейрона
реснички и поэтому способствуют течению спинномозговой жидкости в полостях мозга. Другие развивающиеся спонгиобласты теряют связь как с внутренней, так и с наружной мембранами нервной трубки и становятся астроцитобластами, из которых развиваются астроциты. Клетки, теряющие связь с внутренней пограничной мембраной, называются медуллобластами и дифференцируются в олигодендроциты. Астроциты и олигодендроциты представляют собой два типа глиальных клеток (из трех). Таким образом, почти все клетки нервной ткани имеют общее происхождение (эктодермальное) и дифференцируются в два типа клеток: нейроны и нейроглию. Нейроны Нейроны (рис. 2) являются возбудимыми клетками нервной системы. В отличие от глиальных клеток они способны возбуждаться (генерировать потенциалы действия) и проводить возбуждение. Нейроны высокоспециализированные клетки и в течение жизни не делятся. В нейроне выделяют тело (сому) и отростки. Сома нейрона имеет ядро и клеточные органоиды (рис. 2, 1).Основной функцией сомы является осуществление метаболизма клетки. Рис. 2. Нейрон: 1 — сома (тело) нейрона; 2 — дендрит; 3 — тело Швановской клетки; 4 — миелинизированный аксон; 5 — коллатераль аксона; 6 — терминаль аксона; 7 — аксонный холмик; 8 — синапсы на теле нейрона
Число отростков у нейронов различно, но по строению и выполняемой функции их делят на два типа. Одни — короткие, сильно ветвящиеся отростки, которые называются дендритами (от dendro — дерево, ветвь). Нервная клетка несет на себе от одного до множества дендритов (рис. 2, 2).Основной функцией дендритов является сбор информации от множества других нейронов. Ребенок рождается с ограниченным числом дендритов (межнейронных связей), и увеличение массы мозга, которое происходит на этапах постнатального развития, реализуется за счет увеличения массы дендритов и глиальных элементов. Другим типом отростков нервных клеток являются аксоны (рис. 2, 4).Аксон в нейроне один и представляет собой более или менее длинный отросток, ветвящийся только на дальнем от сомы конце. Эти ветвления аксона называются аксонными терминалами (окончаниями) (рис. 2, 6).Место нейрона, от которого начинается аксон (рис. 2, 7; 6, 2), имеет особое функциональное значение и называется аксонным холмиком (рис. 2, 7). Здесь генерируется потенциал действия — специфический электрический ответ возбудившейся нервной клетки. Функцией же аксона является проведение нервного импульса к аксонным терминалям. По ходу аксона могут образовываться его ответвления — коллатерали (рис.2, 5). В месте отхождения коллатерали (бифуркации) импульс «дублируется» и распространяется как по основному ходу аксона, так и по коллатерали. Часть аксонов центральной нервной системы покрывается специальным электроизолирующим веществом — миелином. Миелинизацию аксонов осуществляют клетки глии. В центральной нервной системе эту роль выполняют олигодендроциты, в периферической — Шванновские клетки (рис. 2, 3; 3, 2), являющиеся Рис. 3. Схема миелинизации аксонов: 1 — связь между телом клетки глии и миелиновой оболочкой; 2 — олигодендроцит; 3 — гребешок; 4 — плазматическая мембрана; 5 — цитоплазма олигодендроцита; 6 — аксон нейрона; 7 — перехват Ранвье; 8 — мезаксон; 9 — петля плазматической мембраны
разновидностью олигодендроцитов. Олигодендроцит оборачивается вокруг аксона, образуя многослойную оболочку. Миелинизации не подвергается область аксонного холмика (рис. 2, 7)итерминали аксона. Цитоплазма глиальной клетки выдавливается из межмембранного пространства в процессе «обертывания». Таким образом, миелиновая оболочка аксона состоит из плотно упакованных, перемежающихся липидных и белковых мембранных слоев (рис. 3, 4).Аксон не сплошь покрыт миелином. В миелиновой оболочке существуют регулярные перерывы — перехваты Ранвье (рис. 3, 7). Ширина такого перехвата от 0,5 до 2, 5 мкм. Функция перехватов Ранвье — быстрое скачкообразное (сальтаторное) распространение потенциалов действия, осуществляющееся без затухания. В центральной нервной системе аксоны различных нейронов, направляющиеся к одной структуре, образуют упорядоченные пучки — Рис. 4. Строение синапса: 1 — аксон пресинаптического нейрона; 2 — микротрубочки; 3 — синаптический пузырек (везикула); 4 — синаптическая щель; 5 — дендрит постсинаптического нейрона; 6 — рецептор для медиатора; 7 — постсинаптическая мембрана; 8 — пресинаптическая мембрана; 9 — митохондрия
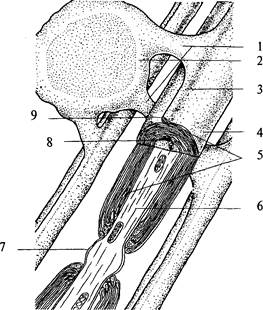
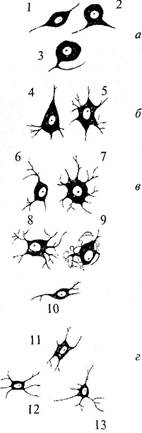
проводящие пути. В подобном проводящем пучке аксоны направляются «параллельным курсом» и часто одна глиальная клетка образует оболочку нескольких аксонов. Поскольку миелин является веществом белого цвета, то проводящие пути нервной системы, состоящие из плотно лежащих миелинизированных аксонов, образуют белое вещество мозга. В сером же веществе мозга локализуются тела клеток, дендриты и немиелинизированные части аксонов. В пределах центральной нервной системы каждая терминаль аксона оканчивается на дендрите, теле или аксоне других нейронов. Контакты между клетками подразделяются в зависимости от того, чем они образованы. Контакт, образуемый аксоном на дендрите, называется аксо-дендритным; аксоном на теле клетки — аксо-соматическим; если он образован двумя аксонами, то называется аксо-аксональным, а двумя дендритами — дендро-дендритным. За пределами ЦНС терминали могут заканчиваться как на нейронных элементах, так и на других возбудимых клетках (мышечных или железистых). В любом случае между нейроном и последующей клеткой образуется специфический контакт — синапс (рис. 4, 4).В образовании синапса участвуют как аксонная терминаль (пресинаптическая часть), так и мембрана последующей клетки (постсинаптическая часть). Синапс состоит из пресинаптической бляшки (расширение терминали аксона), оканчивающейся пресинаптической мембраной (рис. 4, 8), и постсинаптической мембраны (участка мембраны постсинаптической клетки, лежащего под синаптической бляшкой) (рис. 4, 7). Между пресинаптической и постсинаптической мембранами расположена синаптическая щель (рис. 4, 4). От ее величины зависит тип передачи информации через синапс. Если расстояние между мембранами нейронов не превышает 2—4 нм или они контактируют между собой, то такой синапс является электрическим, поскольку подобное соединение обеспечивает низкоомную электрическую связь между этими клетками, позволяющую электрическому потенциалу непосредственно или электротонически передаваться от клетки к клетке. Доля электрических синапсов в ЦНС позвоночных очень мала. Чаще всего мембраны нейронов расположены в непосредственной близости друг к другу и разделены обычным межклеточным пространством (щелью шириной примерно 20 нм) — смежное соединение. Такая смежность мембран облегчает перемещение из одной клетки в межклеточную щель химических веществ (ионов, метаболитов нейронов), которые оказывают влияние как на ту же самую клетку, так и на отростки соседних нейронов. Эти соединения нейронов относят к химическим синапсам. В пресинаптическом окончании химического синапса находятся пузырьки — везикулы (рис. 4, 5), содержащие вещество — передатчик, называемое медиатором. В момент прихода к синаптической бляшке электрического импульса везикулы открываются в пресинаптическую щель, выбрасывая туда медиатор. Медиатор диффундирует через щель и на постсинаптической мембране взаимодействует с рецептором (рис. 4, 6), специфически чувствительным к медиатору, при этом возникает постсинаптический потенциал. Исключением из данного правила являются пептидергические нейроны, не имеющие в пресинаптической области везикул, так как медиатор-пептид синтезируется в соме нейрона и транспортируется по аксону в зону контакта. Таким образом, информация в нервной системе передается только в одном направлении (от пресинаптического нейрона к постсинаптическому) и в этом процессе участвует биологически активное вещество — медиатор. До 50-х годов XX столетия к медиаторам относили две группы низкомолекулярных соединений: амины (ацетилхолин, адреналин, норадреналин, серотонин, дофамин) и аминокислоты (гамма-аминомасляная кислота, глутамат, аспартат, глицин). Позже было показано, что специфическую группу медиаторов составляют нейропептиды, которые могут выступать также и в качестве нейромодуляторов (веществ, изменяющих величину ответа нейрона на стимул). В настоящее время известно, что нейрон может синтезировать и выделять несколько нейромедиаторов (сосуществующие медиаторы). Такое представление о химическом кодировании вошло в основу принципа множественности химических синапсов. Нейроны обладают нейромедиаторной пластичностью, т.е. способны менять основной медиатор в процессе развития. Сочетание медиаторов может быть неодинаковым для разных синапсов. В нервной системе существуют особые нервные клетки — нейросекреторные. Они имеют типичную структурную и функциональную (т.е. способность проводить нервный импульс) нейрональную организацию, а их специфической особенностью является нейросекреторная функция, связанная с секрецией биологически активных веществ. Функциональное значение этого механизма состоит в обеспечении регуляторной химической коммуникации между центральной нервной и эндокринной системами, осуществляемой с помощью нейросекретируемых продуктов. В процессе эволюции клетки, входящие в состав примитивной нервной системы, специализировались в двух направлениях: обеспечение быстро протекающих процессов, т.е. межнейронное взаимодействие, и обеспечение медленно текущих процессов, связанных с продукцией нейрогормонов, действующих на клетки-мишени на расстоянии. В процессе эволюции из клеток, совмещающих сенсорную, проводниковую и секреторную функции, сформировались специализированные нейроны, в том числе и нейросекреторные. Следовательно, нейросекреторные клетки произошли не от нейрона как такового, а от их общего предшественника — пронейроцита беспозвоночных животных. Эволюция нейросекреторных клеток привела к формированию у них, как и у классических нейронов, способности к процессам синаптического возбуждения и торможения, генерации потенциала действия. Для млекопитающих характерны мультиполярные нейросекреторные клетки нейронного типа, имеющие до 5 отростков. Такого типа клетки имеются у всех позвоночных, причем они в основном составляют нейросекреторные центры. Между соседними нейросекреторными клетками обнаружены электротонические щелевые контакты, которые, вероятно, обеспечивают синхронизацию работы одинаковых групп клеток в пределах центра. Аксоны нейросекреторных клеток характеризуются многочисленными расширениями, которые возникают в связи с временным накоплением нейросекрета. Крупные и гигантские расширения называются «телами Геринга». В пределах мозга аксоны нейросекреторных клеток, как правило, лишены миелиновой оболочки. Аксоны нейросекреторных клеток обеспечивают контакты в пре- делах нейросекреторных областей и связаны с различными отделами головного и спинного мозга. Одна из основных функций нейросекреторных клеток — это синтез белков и полипептидов и их дальнейшая секреция. В связи с этим в клетках подобного типа чрезвычайно развит белоксинтезирующий аппарат — это гранулярный эндоплазматический ретикулум и аппарат Гольджи. Сильно развит в нейросекреторных клетках и лизосомальный аппарат, особенно в периоды их интенсивной деятельности. Но самым существенным признаком активной деятельности нейросекреторной клетки является количество элементарных нейросекреторных гранул, видимых в электронном микроскопе. Типы нейронов Конфигурацию отдельного нейрона выявить очень трудно, поскольку они плотно упакованы. Все нейроны принято делить на несколько типов в зависимости от числа и формы, отходящих от их тела отростков. Различают три типа нейронов: униполярные, биполярные и мультиполярные. Униполярные клетки. Клетки, от тела которых отходит только один отросток. На самом деле при выходе из сомы этот отросток разделяется на два: аксон и дендрит. Поэтому правильнее называть их псевдоуниполярными нейронами. Для этих клеток характерна определенная локализация. Они принадлежат неспецифическим сенсорным модальностям (болевая, температурная, тактильная, проприоцептивная) и расположены в сенсорных узлах: спинальных, тройничном, каменистом (рис. 5а). Биполярные клетки — это клетки, которые имеют один аксон и один дендрит. Они характерны для зрительной, слуховой, обонятельной сенсорных систем (см. рис. 5 a). Мультиполярные клетки имеют один аксон и множество дендритов. К такому типу нейронов принадлежит большинство нейронов ЦНС. Исходя из особенностей формы этих клеток их делят на веретенообразные, корзинчатые, звездчатые, пирамидные (рис. 5 б, в, г).Только в коре головного мозга насчитывается до 60 вариантов форм тел нейронов. Сведения о форме нейронов, их местоположении и направлении отростков очень важны, поскольку позволяют понять каче- Рис. 5. Типы нейронов: а — сенсорные нейроны: 1 — биполярный; 2 — псевдобиполярный; 3 — псевдоуниполярный; б — двигательные нейроны: 4 — пирамидная клетка; 5 — мотонейроны спинного мозга; 6 — нейрон двойного ядра; 7 — нейрон ядра подъязычного нерва; в — симпатические нейроны: 8 — нейрон звездчатого ганглия; 9 — нейрон верхнего шейного ганглия; 10 — нейрон бокового рога спинного мозга; г — парасимпатические нейроны: 11 — нейрон узла мышечного сплетения кишечной стенки; 12 — нейрон дорсального ядра блуждающего нерва; 13 — нейрон ресничного узла
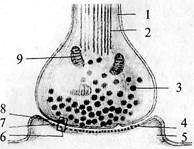
ство и количество связей, приходящих к ним (структура дендритного дерева), и пункты, в которые они посылают свои отростки. Глия Греческое слово «глия» означает «клей». Глиальные клетки впервые описал в 1846 г. Р. Вирхов, который считал, что они «склеивают», скрепляют нервные клетки, «придавая целому его особую форму». Глиальные клетки выполняют в нервной системе множество еще не совсем ясных функций. В отличие от нейронов глиальные клетки сохраняют способность к делению в течение всей жизни. Рис. 6. Схема взаиморасположения элементов нервной ткани: 1 — синапс; 2 — аксонный холмик; 3 — миелиновая оболочка аксона; 4 — астроцит; 5 — тело нейрона; 6 — синаптическая бляшка; 7 — дендрит; 8 — капилляр
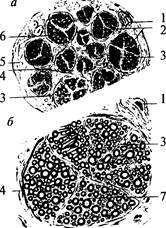
Хотя они имеют мембранный потенциал, но способны генерировать потенциал действия — возбуждаться. Нейроглия составляет почти половину объема мозга, а число клеток глии значительно превышает число нейронов (по меньшей мере в 10 раз). Различают три типа глиальных клеток: астроглия, олигодендроглия и микроглия. Астроглия. Астроглия. Происходит из спонгиобластов, развивающихся в клетки, имеющие множество отростков. Длинные извитые отростки астроцитов переплетаются с отростками нейронов. Значительное число отростков астроцитов представляют собой «ножки», плотно прилегающие к капиллярам и покрывающие собой почти всю поверхность сосуда (рис. 6, 4).Астроциты, расположенные в местах концентрации тел нейронов (серое вещество), образуют Рис. 7. Поперечный срез через нерв (а) и нервный ствол (б): 1 — кровеносный сосуд; 2 — нервные волокна; 3 — эндонервий; 4 — перинервий; 5 — эпинервий; 6 — пучки нервных волокон; 7 — мякотное нервное волокно
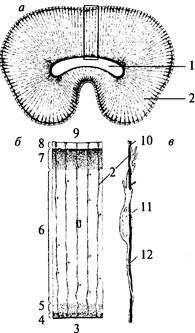
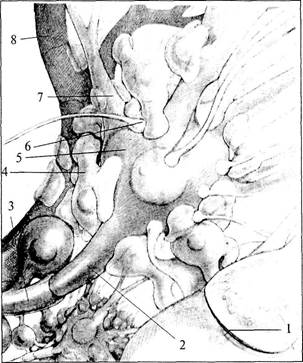
больше отростков, чем астроциты в белом веществе. Таким образом, астроциты — это клетки, располагающиеся между капиллярами и телами нейронов и осуществляющие транспорт веществ из крови в нейроны и обратно. Кроме того, астроглия связывает с кровеносным руслом спинномозговую жидкость. Олигодендроглия. Олигодендроглия. Олигодендроциты имеют то же происхождение, что и астроциты. По размерам они меньше, чем астроциты и имеют меньше отростков. Основная масса олигодендроцитов располагается в белом веществе мозга и ответственна за образование миелина. Эти олигодендроциты обладают длинными отростками. Олигодендроциты, расположенные в периферической нервной системе, называются Шванновскими клетками (см. рис. 2, 3; 3, 2).Те олигодендроциты, которые находятся в сером веществе, располагаются, как правило, вокруг тел нейронов, плотно прилегая к ним. Поэтому их называют клетками-сателлитами. Они характеризуются наличием коротких отростков (см. рис. 6, 3). Микроглия. Микроглия. Клетки микроглии происходят из мезодермы. Как видно из названия, они отличаются небольшими размерами. Эти клетки могут активно передвигаться и выполнять фагоцитарные функции. Благодаря способности к активной миграции микроглия распределена по всей центральной нервной системе. Строение нервов Нервы образованы многочисленными пучками мякотных и безмякотных нервных волокон, которые объединяются в нервные стволы и изолируются соединительной тканью (рис. 7). Наружная оболочка нервного ствола представлена эпинервием — рыхлой неоформленной соединительной тканью, богатой коллагеновыми волокнами, фибробластами, жировыми клетками, а также кровеносными и лимфатическими сосудами. От эпинервия внутрь нервного ствола расположен перинервий, который представляет собой тонкие прослойки соединительной ткани, которые разделяют нерв на пучки. Перинервий также содержит кровеносные и лимфатические сосуды. Соединительная ткань внутри нерва — эндонервий — связывает отдельные нервные волокна в пучки (см. рис. 7).
|
|||||||||
|
|
Последнее изменение этой страницы: 2017-02-17; просмотров: 73; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.21.104.109 (0.111 с.) |