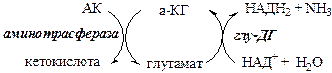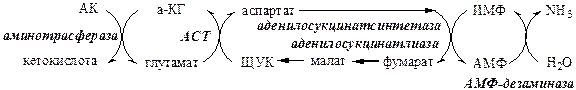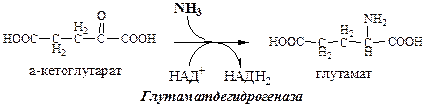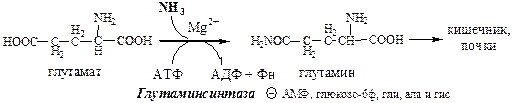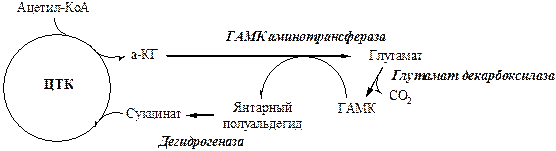Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Тема: Биохимия нервной тканиСтр 1 из 2Следующая ⇒
ЛЕКЦИЯ № 28 Тема: Биохимия нервной ткани Факультеты: лечебно-профилактический, медико-профилактический, педиатрический. 2 курс.
Нервная система: определение понятия Функции нервной системы: 1. Воспринимает информацию из внешней и внутренней среды; 2. Перерабатывает полученную информацию; 3. Хранит полученную информацию; 4. Генерирует сигналы, обеспечивающие ответные реакции, адекватные действующим раздражителям; Благодаря этому, нервная система координирует взаимодействие организма с внешней средой, координирует функции различных органов и тканей и осуществляет интеграцию частей организма в единое целое, является центральным органом поддержания гомеостаза.
КЛАССИФИКАЦИЯ НЕРВНОЙ СИСТЕМЫ Анатомически нервную систему условно подразделяют на: 1. центральную нервную систему (ЦНС), которая включает головной и спинной мозг; 2. периферическую нервную систему (ПНС), к которой относят периферические нервные узлы, нервы и нервные окончания. Физиологически, в зависимости от характера иннервации органов и тканей, нервную систему разделяют на: 1. соматическую (анимальную) нервную систему, которая регулирует преимущественно функции произвольного движения. 2. автономную (вегетативную) нервную систему, которая регулирует деятельность внутренних органов, сосудов и желез. Она осуществляет адаптационно-трофическую функцию. а). симпатическая нервная система (СНС); б). парасимпатическая нервная система (ПСНС). СНС и ПСНС различаются по локализации центров в мозге и периферических узлов, а также характером влияния на внутренние органы. КЛАССИФИКАЦИЯ НЕРВНОЙ ТКАНИ Функциональной тканью нервной системы является нервная. Нервная ткань – это высокоспециализированная ткань, обладающая возбудимостью и проводимостью, она состоит из нейронов и нейроглии (макро- и микроглия). По клеточному составу нервную ткань делят на серое и белое вещество; Серое вещество образованоскоплением нейронов, тонких немиелинизированных нервных волокон и нейроглии (астроциты, олигодендроциты), которое в ЦНС называется ядром, а в ПНС – ганглием (узлом). Белое вещество представлено совокупностью аксонов, покрытых миелиновой оболочкой и глиальных клеток (астроцитов). Такие пучки нервных волокон в ЦНС носят название трактов, в ПНС они образуют нервы. Для каждого тракта, характерно преобладание волокон, образованных однотипными нейронами.
КЛЕТКИ НЕРВНОЙ ТКАНИ Нейрон Нейрон - это функциональная единица нервной системы, он состоит из тела (сомы), многочисленных ветвящихся коротких отростков – дендритов и одного длинного отростка – аксона, длина которого может достигать несколько десятков сантиметров. Аксоны и дендриты оканчиваются синаптическими образованиями. Дендриты, проводят нервный импульс по направлению к телу клетки, а аксон, проводит его от сомы. Таким образом, дендриты и аксоны отвечают соответственно за получение и передачу сигнала. Тело нейрона является трофическим центром, нарушение целостности которого ведет клетку к гибели. Тело нейрона окружено плазматической мембраной – плазмалеммой. Плазмалемма выполняет структурную функцию, служит барьером для поддержания внутриклеточного состава (клеточные органеллы, везикулы нейромедиаторов, метаболиты), играет активную (ионные насосы, ферменты) и пассивную (ионные каналы, высвобождение нейромедиатора) роли в создании мембранного потенциала, транспорте веществ через мембрану и передаче нервного импульса. Внутри нейрон заполнен нейроплазмой (цитоплазмой). Объем нейроплазмы аксона и дендритов, может в несколько раз превышать объем нейроплазмы в теле нейрона. Нейроплазма содержит все основные органеллы клетки. В теле нейрона и проксимальных отрезках дендритов под плазмалеммой находится так называемая подповерхностная мембранная структура. Это - цистерны, которые расположены параллельно поверхности плазмалеммы и отделены от нее очень узкой светлой зоной. Предполагают, что цистерны играют важную роль в метаболизме нейрона. ЭПС нейрона хорошо развита. Мембраны ЭПС связаны с плазмалеммой и оболочкой ядра нейрона. В комплексе Гольджи сосредоточены главным образом липидные компоненты клетки. М итохондрии нейронов содержат меньше ферментов, участвующих в процессах окисления ЖК и АК, чем митохондрии других тканей. Лизосомы в нейроне обнаруживаются постоянно.
В нейроплазме содержатся специальные органоиды – нейрофибриллы и вещество Ниссля (тигроид). Тигроид представляет собой глыбки базофильного вещества, состоящего из РНК и белков, располагающиеся вокруг ядра и заходящие в основания дендритов. Нейрофибриллы – тонкие нити, расположенные в разных направлениях и формирующие густую сеть; они состоят из очень тонких (70 – 200 А) протофибрилл. Нейрофибриллы служат поддерживающим остовом нейрона. Глиальные клетки Нейроглия (от греческого glia – клей) это клетки нервной системы, которые не проводят нервные импульсы. Глиальные клетки занимают 50% объема центральной нервной системы человека и составляют более 90% от всех ее клеток. В ЦНС выделяют 2 вида глии: 1. Макроглия · Астроцитарная глия обеспечивает микроокружение нейронов, выполняет опорную и трофическую функции в сером и белов веществе, участвует в метаболизме нейромедиаторов, входят в состав гематоэнцефалического барьера. · Эпендимная глия образует выстилку желудочков головного мозга и входит в состав гематоликворного барьера. · Олигодендроглия встречается в сером и белом веществе; она обеспечивает барьерную функцию, участвует в формировании миелиновых оболочек нервных волокон, регулирует метаболизм нейронов, захватывает нейромедиаторы. 2. Микроглия – специализированные макрофаги ЦНС. Активизируются при воспалительных и дегенеративных заболеваниях. Выполняют в ЦНС роль антиген-представляющих дендритных клеток.
Клетки макроглии обладают более высоким мембранным потенциалом, чем нейроны, который является чисто калиевым. Глиальные клетки выполняют роль калиевого буфера, они поддерживают внеклеточную концентрацию К+. Между собой глиальные клетки связаны контактными зонами, через которые происходит метаболический обмен. Напротив, нейроны всегда отделены друг от друга щелью (не менее 20нм). Обмен веществами происходит также между глией и аксонами. Глиальные клетки обеспечивают образование нейронами синапсов. Шванновские клетки (глиальные клетки ПНС), подобно олигодендроглии ЦНС, обертываются вокруг аксона и образуют миелин, который электроизолирует аксон и ускоряет проведение импульса. Шванновские клетки участвуют в восстановлении поврежденных нервов, кроме того, после денервации они могут заменять денервированное нервное окончание в мышце и даже выделять медиатор. Белки нервной ткани В головном мозге на белки приходиться 40% сухой массы. В настоящее время выделено более 100 белковых фракций нервной ткани (методами хроматографии, электрофореза и экстракции буферными растворами). В нервной ткани содержатся простые и сложные белки. Простые белки · Нейроальбумины – основные растворимые белки (89-90%) нервной ткани, являются белковым компонентом фосфопротеинов, в свободном состоянии встречаются редко. Легко соединяются с липидами, нуклеиновыми кислотами, углеводами и другими небелковыми компонентами. · Нейроглобулины, содержатся в небольшом количестве (в среднем 5%). · Катионные белки - основные белки (рН 10,5 – 12,0), например, гистоновые. При электрофорезе они движутся к катоду. · Нейросклеропротеины (опорные белки). Например,нейроколлагены, нейроэлластины, нейростромины и др. Они составляют 8-10% от всех простых белков нервной ткани, локализованы в основном в белом веществе головного мозга и ПНС, выполняют структурно-опорную функцию.
Сложные белки Сложные белки нервной ткани представлены: нуклеопротеинами, липопротеинами, протеолипидами, фосфопротеинами, гликопротеинами и т.д. · Гликопротеины – содержат олигосахаридные цепи, которые придают специфические отличия клеточным мембранам. Нейроспецифические гликопротеины участвуют в формировании миелина, в процессах клеточной адгезии, нерорецепции и взаимном узнавании нейронов в онтогенезе и при регенерации. · Протеолипиды – в наибольших количествах содержатся в миелине и в небольших количествах - в синаптических мембранах и синаптических пузырьках. Нейроспецифические белки В цитоплазме нейронов присутствуют кальцийнейрин, белок 14-3-2, белок S-100, белок Р-400. · Белок S-100 (или кислый белок), содержит много глутаминовой и аспарагиновой кислот, гомологичен мышечному тропонину С, находиться в цитоплазме или связан с мембранами. На 85-90% он сосредоточен в нейроглии, и на 10-15% в нейронах. Участвует в развитии нервной системы и ее пластичности. Концентрация S-100 возрастает при обучении животных. · Белок 14-3-2 -кислый белок, который преимущественно локализован в нейронах ЦНС. · Белок Р-400 находится в мозжечке мышей, где, возможно, отвечает за двигательный контроль. К сократительным белкам нейронаотносятся нейротубулин, нейростенин, актиноподобные белки (кинезин и др.). Они обеспечивают ориентацию и подвижность цитоскелета (микротрубочек и нерофиламентов), активный транспорт веществ в нейроне, участвуют в работе синапсов. В нейронах имеются белки, осуществляющие гуморальную регуляцию. Это некоторые гликопротеины гипоталамуса, нейрофизины и подобные им белки. На мембране нейронов расположены нейроспецифические поверхностные антигены (NS1, NS2, L1) с неизвестной функцией и факторы адгезии клеток(N-САМ), важные для развития нервной системы. Нейроспецифические белки участвуют в осуществлении всех функций нервной системы - генерации и проведении нервного импульса, процессах переработки и хранении информации, синаптической передаче, клеточном узнавании, рецепции и др. Ферменты нервной ткани В мозговой ткани содержится большое количество ферментов, катализирующих обмен белков, жиров и углеводов. Также цитоплазма нейронов содержит ферменты метаболизма посредников и медиаторов.
Мозговая ткань характеризуется высокой активностью: ЛДГ (ЛДГ1,ЛДГ2), АСТ, альдолазы, креатинкиназы (ВВ), гексокиназы, малатдегидрогеназы, глутаматдегидрогеназы, холинэстеразы, кислой фосфатазы, моноаминоксидазы. В глиальных клетках преобладает ЛДГ5, а в нейронах - ЛДГ1. Для мозга характерна так же высокая активность ферментов метаболизма циклических нуклеотидов, которые принимают участие в синаптической передаче нервного импульса. Аминокислоты нервной ткани Аминокислотный фонд мозга человека составляет в среднем 34ммоль на 1г ткани, что значительно превышает их содержание, как в плазме крови, так и в СМЖ. Высокая концентрация АК в нервной ткани достигается путем их многоступенчатого активного и пассивного транспорта из плазмы крови. Сначала АК переносятся через гематоэнцефалический барьер эндотелия мозговых капилляров, затем они переходят из внеклеточной жидкости в клетки мозга, а далее – в субклеточные органеллы. Активность систем транспорта аминокислот, так же как и состав их пула, изменяются в процессе развития мозга. Аминокислоты проникают в мозг молодых животных быстрее и достигают более высоких концентраций, чем у взрослых. Более 50% α-аминоазота головного мозга приходится на долю глутаминовой кислоты, глутамина и глутатиона. Специфичными для мозговой ткани являются ГАМК, N-ацетиласпарагиновая кислота и цистатионин. В нервной ткани аминокислоты распределяются неравномерно, в основном это касается аминокислот, выполняющих функцию нейромедиаторов (глутаминовая кислота, ГАМК, глицин и др). N-ацетиласпарагиновой кислоты больше в сером веществе, чем в белом. Также она есть в ПНС и сетчатке. Содержание цистаниона выше в белом веществе, чем в сером, и оно повышается в процессе развития.
Содержание (мкмоль/г) свободных аминокислот в мозге, плазме и СМЖ человека
Липиды нервной ткани Нервная ткань отличается высоким содержанием и разнообразием липидов, которые придают ей специфические особенности. В сером веществе фосфоглицериды составляют более 60% от всех липидов, а в белом – около 40%. В белом веществе содержится больше холестерина, сфингомиелинов и особенно цереброзидов, чем в сером веществе.
· Холестерин составляет около 25% от общего содержания липидов. При этом в мозге мало эфиров холестерина. ХС повышает электроизоляционные свойства клеточных мембран, защищает их от ПОЛ, повышает их механическую прочность. · Свободных жирных кислот в мозге мало, а э терефицированных жирных кислот в мозге очень много, в основном это пальмитиновая, стеариновая, олеиновая и арахидоновая кислоты. · Сфинголипиды (ганглиозиды и цереброзиды) участвуют в процессах коммуникации нервной клетки с окружающей ее средой. Они обеспечивают передачу сигналов с наружной поверхности клетки в ее внутреннее пространство. Разнообразие углеводных частей сфинголипидов делает их носителями специфической информации. · Ганглиозиды находятся преимущественно в сером веществе. Выделяют 4 основных ганглиозида – Gм1, GD1a, GD1b и GT1. Зрительная кора относительно богата GT1 и GD1b, в коре головного мозга содержатся в основном GT1, в белом веществе - Gм1. Функции ганглиозидов: 1). являются рецепторами внешних сигналов; 2). с гликопротеинами отвечают за специфичность клеточной поверхности, распознавание клеток и их адгезию; 3). участвуют в развитии нервной системы при образовании «правильных» межклеточных связей; 4). участвуют в коммуникации между мембранами аксонов и окружающими их олигодендроглиальными клетками; 5). участвуют в функциональной адаптации зрелой нервной системы. Синтез ганглиозидов связан с дифференциацией нейронов. · Цереброзиды находятся преимущественно в белом веществе, особенно их много миелине. В цереброзидах и сульфатидах ЦНС превалируют производные галактозы. Количество галактоцерамидов и галактосульфатидов возрастает по мере миелинизации. · Фосфатидилинозитолы в ЦНС не превышают 0,5-2% от общих липидов. Локализованы в плазматических мембранах, в миелине (до 95% всех фосфоинозитов мозга), в ЭПС, наружной митохондриальной и ядерной мембранах. Участвуют в инозитолтрифосфатной системе передаче сигнала с мускариновых и α1-адренергических рецепторов. Углеводы нервной ткани По сравнению с другими тканями ткань мозга содержит мало глюкозы и гликогена. У новорожденных концентрация гликогена в мозге выше, чем у взрослых. Олигосахариды составляют 2-10% массы плазматической мембраны, большая их часть связана с белками и меньшая с - гликолипидами. Практически все они локализованы на внешней поверхности плазматической мембраны и придают ей индивидуальность и специфичность. Нуклеотиды нервной ткани Большинство нейронов ЦНС диплоидны, а небольшая их часть в некоторых отделах ЦНС (клетки Пуркинье мозжечка) может содержать избыточное количество ДНК. Особенностями хроматина нейронов являются необычно короткие нуклеосомные единицы, наличие редких вариантов гистонов, большое разнообразие негистоновых белков и высокая матричная активность. Содержание РНК в нейронах велико, что связано с активным синтезом белка. Среднее отношение РНК/ДНК может достигать 50 и редко бывает ниже 3. В печени, поджелудочной железы, почках оно составляет 2-4,5. Содержание цАМФ и цГМФ в головном мозге значительно выше, чем во многих других тканях. Уровень цАМФ в мозге составляет в среднем 1-2, а цГМФ – до 0,2 нмоль на 1г ткани. Безмиелиновое волокно Безмиелиновое волокно состоит из 7-12 тонких аксонов, которые проходят внутри тяжа, образованного цепочкой нейроглиальных клеток. Безмиелиновые волокна имеют постганглионарные нервные волокна, входящие в состав вегетативной нервной системы. Миелиновое волокно Миелиновое волокно состоит из одного аксона, который окутан миелиновой оболочкой и окружен глиальными клетками. Миелиновая оболочка образована плазматической мембраной Шванновской или олигодендроглиальной клетки, которая сложена вдвое и многократно обернута вокруг аксона. По длине аксона миелиновая оболочка образует короткие чехольчики - междоузлия, между которыми имеются немиелизированные участки – перехваты Ранвье. Миелиновое волокно более совершенно, чем безмиелиновое, т.к. оно обладает более высокой скоростью передачи нервного импульса. Миелиновые волокна имеют проводниковая система соматической нервной системы, преганглионарные волокна вегетативной нервной системы.
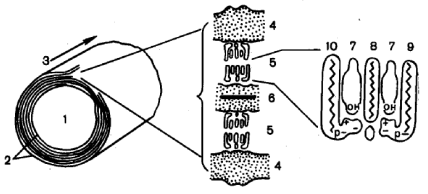
Молекулярная организация миелиновой оболочки (по Х.Хидену) 1-аксона; 2-миелин; 3-ось волокна; 4-белок (наружные слои); 5-липиды; 6-белок (внутренний слой); 7-холестерин; 8-цереброзид; 9- сфингомиелин; 10-фосфатидилсерин. Химический состав миелина Миелин содержит много липидов, мало цитоплазмы и белков. Мембрана миелиновой оболочки в расчете на сухую массу содержит 70% липидов (что в целом составляет около 65% всех липидов мозга) и 30% белков. 90% всех липидов миелина приходиться на холестерин, фосфолипиды и цереброзиды. Миелин содержит немного ганглиозидов. Белковый состав миелина периферической и центральной нервной системы различен. Миелин ЦНС содержит три белка: 1. Протеолипид, составляет 35 – 50% от общего содержания белка в миелине, имеет молекулярную массу 25кДа, растворим в органических растворителях; 2. Основной белок А1, составляет 30% от общего содержания белка в миелине, имеет молекулярную массу 18кДа, растворим в слабых кислотах; 3. Белки Вольфграма - несколько кислых белков большой массы растворимых в органических растворителях, функция которых неизвестна. Составляют 20% от общего содержания белка в миелине. В миелине ПНС, протеолипид отсутствует, основной белок представлен белками А1 (немного), Р0 и Р2. В миелине обнаружена ферментативная активность: 1. холестеролэстеразы; 2. фосфодиэстеразы, гидролизирующей цAMФ; 3. протеинкиназы А, фосфорилирующей основной белок; 4. сфингомиелиназы; 5. карбоангидразы. Миелин благодаря своему строению обладает более высокой стабильностью (устойчивостью к разложению), чем другие плазматических мембран.
Серосодержащие аминокислоты Метионин является источником метильных групп и на 80% используется для синтеза белка. Цистатионин важен для синтеза сульфитидов и сульфатилрованных мукополисахаридов.
Обмен азота нервной ткани Непосредственным источником аммиака в головном мозге служит непрямое дезаминирование аминокислот с участием глутаматдегидрогеназы, а так же дезаминирование с участием АМФ–ИМФ цикла.
Обезвреживание токсичного аммиака в нервной ткани происходит с участием α-кетоглутарата и глутамата.
Нервный импульс В отличие от потенциала покоя, потенциал действия охватывает лишь очень небольшой участок аксона (в миелинизированных волокнах – от одного перехвата Ранвье до соседнего). Возникнув в одном участке аксона, потенциал действия вследствие диффузии ионов из этого участка вдоль волокна снижает потенциал покоя в соседнем участке и вызывает здесь то же развитие потенциала действия. Благодаря этому механизму потенциал действия распространяется по нервным волокнам и называется нервным импульсом. В миелинизированном нервном волокне натриевые и калиевые ионные каналы расположены в немиелинизированных участках перехватов Ранвье, где мембрана аксона контактирует с межклеточной жидкостью. Вследствие этого нервный импульс перемещается «скачками»: ионы Na+, поступающие внутрь аксона при открытии каналов в одном перехвате, диффундируют вдоль аксона по градиенту потенциалов до следующего перехвата, снижают здесь потенциал до пороговых значений и тем самым индуцируют потенциал действия. Благодаря такому устройству скорость поведения импульса в миелинизированном волокне в 5-6 раз больше, чем в немиелинизированных волокнах, где ионные каналы расположены равномерно по всей длине волокна и потенциал действия перемещается не скачками, а плавно.
Классификация синапсов 1. По локализации: центральные (ЦНС) и периферические (нервно-мышечные, нейросекреторные синапсы вегетативной НС). 2. По развитию в онтогенезе: стабильные (безусловный рефлекс) и динамические (условный рефлекс) синапсы. 3. По конечному эффекту: тормозные и возбуждающие. 4. По механизму передачи сигнала: электрические, химические и смешанные. Химические синапсы делят: а). по форме контакта: терминальные (колбообразное соединение) и переходящие (варикозное расширение аксона). б). по природе медиатора: холинергические (медиатор ацетилхолин), адренергические (норадреналин), дофаминергические (дофамин), ГАМК-ергические (ГАМК), глициергические (глицин), глутаматергические (глутамат), аспартатергические (аспартат), пептидергические (пептиды), пуринергические (АТФ). Электрические синапсы осуществляют передачу сигнала путем прямого прохождения потенциалов действия. Электрические синапсы сравнительно редки, их роль в ЦНС пока неясна. Передача сигнала между нейронами идет через щелевые контакты (щель около 2нм) с ионными мостиками-каналами. В противоположность химическому синапсу, сигнал через электрический синапс передается быстро и сразу в два направления. Химический синапс осуществляет передачу сигнала с помощью специальных молекул – нейромедиаторов. Нейромедиатор - это соединение, которое синтезируется и запасается в нейроне, высвобождается при проведении нервного импульса и специфически связывается постсинаптической мембраной, где оно активирует или ингибирует постсинаптическую клетку посредством деполяризации и гиперполяризации. Химический синапс состоит 1). из пресинаптического элемента, который ограничен пресинаптической мембраной. Пресинаптический элемент содержит митохондрии и особые пузырьки – синаптические везикулы, в которых хранится медиатор; 2). постсинаптического элемента, который ограничен постсинаптической мембраной. Постсинаптическая мембрана содержит рецепторы к медиатору; 3). внесинаптической области; 4). синаптической щели (толщина 50 нм), заполненной базальной мембраной.
Адренэргнические синапсы Адренэргические синапсы используют в качестве медиаторов катехоламины – норадреналин (НА), дофамин. Адренэргические синапсы находятся в головном мозге и в СНС – в окончаниях постганглионарных волокон. Они возбуждают сердечную мышцу, тормозят ГМК ЖКТ, бронхов. Катехоламины синтезируются в синапсе из тирозина, который образуется из фенилаланина, либо поступает с пищей. Синтезируемые катехоламины запасаются в везикулах. Кроме катехоламинов, везикулы содержат АТФ (в меньших количествах АДФ и АМФ) и ионы кальция. Под действием нервного импульса везикулы сливаются с пресинаптической мембраной, а катехоламины высвобождаются в синаптическую щель. На поссинаптической мембране катехоламины связываются с рецепторами. Существует 4 вида рецепторов к норадреналину: α-1-АР, α-2-АР, β-1-АР, β-2-АР. Под действием норадреналина: α-1-АР (ГМК сосудов кожи, ЖКТ, нейроны головного мозга) и β-1-АР (миокард) деполяризует постсинаптической мембрану и возбуждает эффекторную клетку, α-2-АР пресинаптической мембраны тормозит выделение синапсом следующей порции НА, β-2-АР (ГМК сосудов скелетных мышц, бронхов, матки, коронарных сосудов) поляризует постсинаптической мембрану и тормозит эффекторную клетку. Адренэргический синапс использует свой медиатор экономично: большая часть (80%) катехоламинов реабсорбируются из синаптической щели пресинаптической мембраной и снова упаковываются в везикулы. Часть катехоламинов диффундирует в кровь, часть захватывается эффекторной клеткой и подвергается катаболизму - аминогруппа удаляется окислительным дезаминированием на митохондриальной мембране с участием моноаминооксидазы, а одна из гидроксильных групп ароматического кольца метилируется катехол-О-метилтрансферазой. На работу адренергических синапсов влияют различные вещества: · Резерпин, октадин нарушают депонирование НА в везикулах; · Ипразид ингибирует МАО; · Пирогаллол ингибирует катехол-О-метилтрансферазу. · Эфедрин усиливает выделение НА; · Октадин, орнид ингибируют выделение НА; · Резерпин, кокаин ингибируют захват НА; Болезнь Паркинсона Болезнь Паркинсона – это прогрессирующее, часто фатальное нарушение центральной нервной системы, которое характеризуется ригидностью мышц, затруднениями движения и тремором. Больных узнают по застывшему выражению лица, и слегка согнутой окоченелой позе и замедленными движениями. У некоторых больных основным симптомом является дрожание конечностей. Это заболевание связано с дегенеративным изменением в отделе ЦНС, ответственного за двигательный контроль. Обычно оно наступает во второй половине жизни, а его причины и факторы, провоцирующие болезнь, в основном неизвестны. Заболевание поражает главным образом черное вещество и полосатое тело мозга – области богатые дофамином. У больных концентрация дофамина, продуктов его деградации и ферментов его биосинтеза снижены. В качестве терапии таким больным назначают оральное применение ДОФА предшественника дофамина. Только небольшая доза введенного ДОФА достигает места своего действия, поэтому ежедневные дозы этого препарата достигают 2 –15г. Дозы могут быть значительно снижены, если вводить ДОФА вместе с ингибитором декарбоксилазы, блокирующим его периферическое разложение. Холинэргические синапсы Холинэргические синапсы - это группа различных синапсов использующих ацетилхолин в качестве нейромедиатора. Они играют важную роль в центральной нервной системе, участвуя в таких процессах, как поведение, сознание, эмоции, обучение и память. Ацетилхолин - важный медиатор вегетативной нервной системы, он присутствует во всех ганглиях СНС и ПСНС, в постганглионарных нервных волокнах всех парасимпатических синапсов и некоторых симпатических. К группе холинэргических синапсов относятся нервно-мышечные соединения, образуемые моторными нейронами. Синапсы преганглионарного типа стимулируются никотином и ацетилхолином, поэтому их называют никотиновыми холинэргическими синапсами. Синапсы постганглионарного типа стимулируются мускарином и ацетилхолином, поэтому их называют мускариновыми холинэргическими синапсами. Никотиновые рецепторы содержатся в нервно-мышечных синапсах скелетных мышц и в вегетативных ганглиях; мускариновые в гладких мышцах и мозге. Серотонинэргические синапсы Серотонинэргические синапсы использую в качестве медиатора серотонин, они имеются в различных отделах головного мозга (мозговом стволе, варолиевом мосту, ядрах шва). Серотонин образуется из триптофана путем гидроксилирования в 5 положении и последующего декарбоксилирования. Затем серотонин упаковывается в секреторные гранулы, и под действием потенциала действия высвобождается. Рецепторы серотонина бывают 2 типов М и Д, которые имеют не менее 15 подтипов. Рецепторы к серотонину действуют через аденилатциклазную систему, инозитолтрифосфатную систему и ионные каналы. Инактивация серотонина: · при его окислительном дезаминирования МАО. · при N-ацетилировании с образованием N-ацетилсеротонин. При О-метилирование N-ацетилсеротонина приводит к образованию мелатонина – гормона шишковидной железы. Серотонин играет важную роль в регуляции эмоционального поведения, двигательной активности, пищевого поведения, сна, терморегуляции, участвует в контроле нейроэндокринных систем. Аминокислотные медиаторы Аминокислотные медиаторы подразделяются на две группы: · возбуждающие кислые (глутамат и аспартат) · ингибиторные нейтральные (ГАМК, глицин, β-аланин и таурин). ГАМК ГАМК ингибиторный медиатор. Он содержится в сером веществе головного мозга, в клетках Пуркинье мозжечка, многих ингибиторных промежуточных нейронов, например, полосатого тела, спинного мозга и коры. ГАМК образуется и разрушается в ГАМК-шунте ЦТК.
Ингибирование заключается в том, что он открывает хлорные каналы, вызывает гиперполяризацию и тормозит возбудимость постсинаптической мембраны эффекторной клетки. Если ингибирующее действие ГАМК-эргических нейронов снимается, то это приводит к неконтролируемой активности связанных с этим медиатором нервных связей. Антагонисты ГАМК, например пикротоксин и бикукуллин, являются, следовательно, мощными конвульсантами. Вещества, усиливающие ингибиторный эффект ГАМК, являются релаксантами и транквилизаторами. На работу ГАМК-реактивных синапсов влияют различные вещества: · Производные гидразина ингибируют синтез ГАМК. · Антогонисты ГАМК: бициклофосфаты, норборнан. · Пресинапсические блокаторы высвобождения ГАМК: тетанотоксин.
Глицин Глицин - основной ингибиторный медиатор спинного мозга и ствола головного мозга. Он открывает хлорные каналы, вызывает гиперполяризацию и тормозит возбудимость постсинаптической мембраны. Глутамат Глутамат - основной возбуждающий медиатор ЦНС. Он представлен в высокой концентрации в нервной ткани (10 мМ) (причем в нейронах выше, чем в глии). Непосредственный источник глутамата в мозговой ткани – восстановительное аминирование и переаминирования α-кетоглутаровой кислоты. Выделено пять рецепторов глутамата. NMDA, АМРА и каинатные рецепторы связаны с Са2+-каналами. Под действием глутамата, рецепторы открывают Са2+-каналы и запускают Са2+ из межклеточного пространства в в нейроплазму. ACPD – рецепторы активируют инозитолтрифосфатную систему. Под действием глутамата они выпускают Са2+ из ЭПС в в нейроплазму. Активацие L-AP4-рецепторов приводит к усилению гидролиза цГМФ и блокаде входящих ионных токов. Глутамат играет важную роль в осуществлении пластичности синапсов и эксайтотоксичности, участвует в развитии долговременной потенциации - процесса, который лежит в основе некоторых форм обучения. Вещество Р Вещество Р – нейромедиатор пресинаптических окончаний С-волокон первичных сенсорных нейронов, образующих синапсы на сенсорных нейронов второго порядка в задних рогах спинного мозга. Он участвует в восприятии болевых сигналов. ХИМИЧЕСКИЕ ОСНОВЫ БОЛИ В основе любой боли лежит раздражение болевых или полимодальных рецепторов, т.е. боль - прежде всего ощущение. Но, поскольку это ощущение влечет за собой стимуляцию различных отделов нервной и эндокринной системы, боль как явление у человека включает эмоциональный, вегетативный, двигательный и поведенческий компоненты, а значит, представляет собой основанное на болевом ощущении психофизиологическое состояние организма. В шестидесятые годы нашего столетия было обнаружено, что два вида болевой чувствительности имеют различный проводниково-рецепторный аппарат и центральные адреса [9]. Болевые рецепторы Поверхностные ткани снабжены нервными окончаниями различных афферентных волокон. Наиболее толстые, миелинизированные Аβ-волокна обладают тактильной чувствительностью. Они возбуждаются при неболезненных прикосновениях и при перемещении. Эти окончания могут служить как полимодальные неспецифические болевые рецепторы только при патологических условиях, например, вследствие возрастания их чувствительности (сенсибилизации) медиаторами воспаления. Слабое раздражение полимодальных неспецифических тактильных рецепторов приводит к чувству зуда. Порог их возбудимости понижают гистамин и серотонин. Специфическими первичными болевыми рецепторами (нонирецепторами) служат два других типа нервных окончаний – тонкие миелинизированные Аδ-терминали и тонкие немиелинизированные С-волокна, филогенетически более примитивны. Оба эти типа терминалей представлены и в поверхностных тканях, и во внутренних органах. Ноцирецепторы дают чувство боли в ответ на самые разные интенси
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-02-07; просмотров: 303; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.220.137.164 (0.124 с.) |