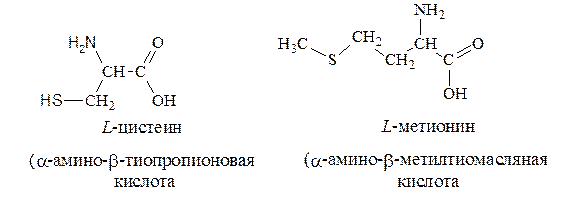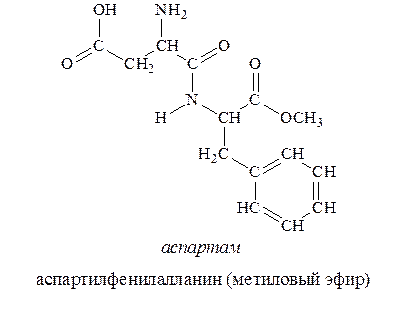Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Строение и общие свойства аминокислот.Стр 1 из 6Следующая ⇒
БИОЛОГИЧЕСКАЯ ХИМИЯ КОНСПЕКТ ЛЕКЦИЙ для студентов специальностей
БЕЛКИ
Могилев, 2004 УКД ББК Рассмотрен и рекомендован к изданию кафедрой Протокол № __ от «__»_____________2004 г. Рассмотрен и рекомендован к изданию секцией выпускающих кафедр. Протокол № __ от «__»_____________2004 г. Составители: доцент Макасеева О.Н. Рецензент: доцент Шуляк Т.Л. Ó Могилевский государственный университет продовольствия Содержание 1 БЕЛКИ.. 4 1.1 Строение и общие свойства аминокислот. 4 1.2 Классификация аминокислот. 7 1.3 Кислотно-основные свойства аминокислот. 11 1.4 Спектры поглощения аминокислот. 14 1.5 Химические реакции аминокислот. 14 1.6 Пептиды. 15 1.7 Белки. 19 1.8 Строение белковой молекулы. 20 1.9 Физико-химические свойства белков. 28 1.10 Выделение белков и установление их однородности. 33 1.11 Классификация белков. 35 2 НУКЛЕИНОВЫЕ КИСЛОТЫ. 38 2.1 Состав нуклеиновых кислот. 38 2.2 Нуклеозиды. 41 2.3 Нуклеотиды.. 42 2.4 Первичная структура нуклеиновых кислот. 45 2.5 Вторичная и третичная структуры ДНК. 46 2.6 Структура РНК.. 49 Рекомендуемая литература. 50 БЕЛКИ Классификация аминокислот. Существует несколько видов классификаций аминокислот входящих в состав белка. В основу одной из классификаций положено химическое строение радикалов аминокислот. Различают аминокислоты: 1. Алифатические – глицин, аланин, валин, лейцин, изолейцин:
2. Гидроксилсодержащие – серин, треонин:
3. Серосодержащие – цистеин, метионин:
4. Ароматические – фенилаланин, тирозин, триптофан:
5. С анионобразующими группами в боковых цепях-аспарагиновая и глутаминовая кислоты:
6. амиды аспарагиновой и глутаминовой кислот – аспарагин, глутамин.
7. Основные – аргинин, гистидин, лизин.
8. Иминокислота – пролин.
Второй вид классификации основан на полярности R-групп аминокислот. Различают полярные и неполярные аминокислоты. У неполярных в радикале есть неполярные связи С–С, С–Н, таких аминокислот восемь: аланин, валин, лейцин, изолейцин, метионин, фенилаланин, триптофан, пролин. Все остальные аминокислоты относятся к полярным (в R-группе есть полярные связи С–О, С–N, –ОН, S–H). Чем больше в белке аминокислот с полярными группами, тем выше его реакционная способность. От реакционной способности во многом зависят функции белка. Особенно большим числом полярных групп, характеризуются ферменты. И наоборот, их очень мало в таком белке как кератин (волосы, ногти).
Аминокислоты классифицируют и на основе ионных свойств R-групп (таблица 1). Кислые (при рН=7 R-группа может нести отрицательный заряд) это аспарагиновая, глутаминовая кислоты, цистеин и тирозин. Основные(при рН =7 Все остальные аминокислоты относятся к нейтральным (группа R незаряжена).
Таблица 1 – Классификация аминокислот на основе полярности
По числу амино и карбоксильных групп аминокислоты делятся на моноаминомонокарбоновые, содержащие по одной карбоксильной и аминогруппа; моноаминодикарбоновые (две карбоксильные и одна аминогруппа); диаминомонокарбоновые (две аминные и одна карбоксильная группа). По способности синтезироваться в организме человека и животных все аминокислоты делятся на заменимые, незаменимые и частично незаменимые.
Незаменимые аминокислоты не могут синтезироваться в организме человека и животных они обязательно должны поступать вместе с пищей. Абсолютно незаменимых аминокислот восемь: валин, лейцин, изолейцин, треонин, триптофан, метионин, лизин, фенилаланин. Частично незаменимые синтезируются в организме, но в недостаточном количестве, поэтому частично должны поступать с пищей. Такими аминокислотами являются аргинин, гистидин, тирозин. Заменимые аминокислоты синтезируются в организме человека в достаточном количестве из других соединений. Растения могут синтезировать все аминокислоты. Пептиды. Аминокислоты соединяются друг с другом ковалентной пептидной связью. Образование ее происходит за счет a-аминогруппы (–NH2) одной аминокислоты и a-карбоксильной (–СООН) группы другой с выделением молекулы воды.
В результате реакции поликонденсации можно получить соединения, составленные из многих аминокислотных остатков – полипептиды. При написании формулы линейных пептидов с известной последовательностью аминокислотных остатков начинают с N-конца (на конце пептида находится свободная a-аминогруппа), используя сокращенные обозначения аминокислот. Названия пептидов складываются из названий соответствующих аминокислот с суффиксом – ил, начиная с N-концевого остатка, – название С-концевой аминокислоты (содержит свободную a-карбоксильную группу) сохраняется. Например, аргинил-аланил-глицин-глутамил-лизин. Каждый пептид содержит только одну свободную a-амино- и Гидролиз пептидных связей может быть осуществлен также действием некоторых ферментов, которые расщепляют пептидные связи избирательно, с образованием коротких пептидов. Например трипсин гидролизует связи образованные карбоксильными группами лизина, аргинина; химотрипсин-карбоксильными группами фенилаланина, тирозина, триптофана. Такой избирательный анализ оказывается очень полезным при установлении аминокислотной последовательности белков и пептидов. Кроме пептидов, образующихся в результате частичного гидролиза молекул белка, существует много пептидов, встречающихся в живых организмах как свободные соединения. Многие природные пептиды отличаются по своей структуре от белков; такие пептиды имеются во всех типах организмов. В структурном отношении пептиды небелковой природы весьма разнообразны: отличаются по размерам, наличию циклических структур, разветвленности, наличию D- и
Карнозин и Ансерин. Эти дипептиды найдены в мышечных тканях позвоночных, в том числе и в мышцах человека. Оба они содержат b-аланин – структурный изомер a-аланина.
Эти дипептиды служат для поддержания постоянного рН в клетках мышц, т.е действуют как буферы, также они участвуют в сокращении мышц, в процессах окислительного фосфорилирования т.е в образовании АТФ. Глутатион (g-глутамилцистеинилглицин) – трипептид, присутствует во всех животных, растениях и микроорганизмах.
Отличительная структурная особенность глутатиона, состоит в том, что глутаминовая кислота в составе этого пептида представляет для образования пептидной связи g-карбоксильную (а не a-карбоксильную группу). Существуют две формы глутатиона восстановленная (SH-глутатион) и окисленная (S-S-глутатион). Взаимопревращения одной формы в другую катализируется ферментом глутатионредуктазой.
В настоящее время известны лишь некоторые из физиологических функций глутатиона: 1) участие в транспорте аминокислот через клеточные мембраны; 2) поддержание восстановленного состояния железа (Fe+2) в гемоглабине; 3) входит в состав фермента глутатионпероксидазы, который защищает клетки от разрушающего действия Н2О2. 4) участвует в детоксикации ряда чужеродных для живой клетки соединений (галогенсодержащие алифатические или ароматические углеводороды) переводит их в водорастворимые соединения, которые выводятся из организма почками. 5) восстановленный глутатион защищает SH-группы белка от окисления, сам при этом превращается в окисленный. Глутатион влияет и на технологические свойства зерна и муки. Восстановленный глутатион вызывает восстановление и разрыв дисульфидных связей в молекуле белков клейковины, т.е разрушает ее. Тесто из такой муки обладает плохими структурно-механическими свойствами), оно ослабляется, расплывается из него нельзя получить хлеб нормального качества.) Много глутатиона в старых дрожжах и зародышах зерновых, что следует учитывать в хлебопечении. Восстановленный глутатион способен активировать протеиназы (ферменты ращепляющие белки) зерна и муки, при этом начинается усиленно протекать протеолиз белков клейковины и вызванное им разжижение теста. Глутатион способствует накоплению в пиве азотистых соединений сравнительно большой молекулярной массы, что вызывает образование мути в пиве и ухудшает его потребительские свойства.
С 1981г. разрешено использовать в качестве низкокалорийной добавки для придания продуктам сладкого вкуса аспартам (торговое название). Аспартам в 200 раз слаще сахара и представляет собой метиловый эфир дипептида, состоящего из остатков аспарагиновой кислоты и фенилаланина.
У млекопитающих (в том числе у человека) вырабатываются пептиды обладающие гормональным регуляторным действием, причем диапазон приложения их действия и эффективность в организме очень разнообразны. Например, два циклических нонапептида вырабатывает гипофиз. Окситоцин стимулирует сокращение матки у беременных самок и выделение молока у кормящих самок. Вазопрессин обладает сильным антидиуретическим действием и участвует в контроле кровяного давления. Соматостатин – один из гормонов гипоталамуса – ингибирует синтез гормона роста человека в гипофизе, что приводит к задержке роста и развития тела. В 1975г. открыта группа пептидов, которые оказывают влияние на передачу нервных импульсов. Их также называют опиатными пептидами, поскольку механизм их действия сходен с механизмом действия морфина и других опиоидов. Они присутствуют в очень малых количествах, как у позвоночных, так и у беспозвоночных. Эти вещества обладают сильным обезболивающим действием, а также участвуют в регуляции настроения и поведения. Белки. Полипептиды, содержащие больше 51 аминокислоты, относятся к белкам. Белки входят в состав всех клеток и тканей живых организмов. Около 50% сухого вещества клетки приходится на белки. Белки характеризуются определенным элементарным составом. Химический анализ показал наличие во всех белках углерода (50-55%), кислорода (21-24%), азота (15-18%), водорода (6-7%), серы (0,3-2,5%). В составе отдельных белков обнаружены также фосфор, йод, железо, медь и некоторые другие макро и микроэлементы, в различных, часто очень малых количествах. Белками (протеинами, от греческого protas – первый, важнейший) называют высокомолекулярные природные полимеры, молекулы которых построены из остатков аминокислот. Поразительно то, что все белки во всех организмах построены их одного и того же набора – 20-ти аминокислот, каждая из которых не обладает никакой биологической активностью. Что же тогда придает белку специфическую активность, одним ферментативную, другим, гормональную, третьим защитную и т.д. Ответ довольно прост: белки отличаются друг от друга тем, что каждый имеет свою характерную для него аминокислотную последовательность. Аминокислоты – это алфавит белковой структуры; соединив их в различном порядке можно получить бесконечное число последовательностей, а, следовательно, и бесконечное количество разнообразных белков, выполняющих различные биологические функции.
1. Ферментативная (каталитическая). В биологических системах почти все реакции катализируются специфическими белками – ферментами. В настоящее время открыто около 300 различных ферментов, каждый из которых служит катализатором определенной биологической реакции. Синтез и распад веществ, их регуляция, перенос химических групп и электронов от одного вещества к другому осуществляется с помощью ферментов. 2. Строительная, структурная функция. Белки образуют основу протоплазмы любой живой клетки, в комплексе с липидами они являются основным структурным материалом всех клеточных мембран всех органелл. 3. Двигательная функция. Любые формы движения в живой природе (работа мышц, движение ресничек и жгутиков у простейших, движение протоплазмы в клетке и т.д.) осуществляется белковыми структурами. 4. Транспортная функция. Перенос различных молекул, ионов осуществляется специфическими белками. Например, белок крови гемоглобин переносит кислород к тканям. Перенос жирных кислот по организму происходит с участием другого белка крови-альбумина. 5. Регуляторная функция. Регуляция углеводного, белкового, липидного обменов осуществляется с помощью гормонов, которые по своему строению относятся к белкам (инсулин) или пептидам (окситоцин, вазопрессин и др.). 6. Защитная – эту функцию выполняют иммуноглобулины (антитела). Они обладают способностью обезвреживать бактерии, вирусы, чужеродные белки, попавшие в организм извне. Процесс свертывания крови, защищающий организм от ее потери, основан на превращениях белка – фибриногена. Кератин – белок волосяного защитного покрова. 7. Фоторецепторные белки: например, родопсин, участвующий в зрительных процессах. 8. Резервные белки используются, как запасной материал для питания развивающегося зародыша и новорожденного организма – это белки семян зернобобовых культур, альбумин – яичный белок, казеин молока. Ферретин – белок животных тканей в котором запасено железо. Резервные белки являются важнейшими компонентами растительной и животной пищи. Имеется много других белков, функции которых довольно необычны. Например, монеллин – белок, выделенный из африканского растения, имеет очень сладкий вкус. Его изучают как вещество нетоксичное и не способствующее ожирению, с целью использования в пищу вместо сахара. Плазма крови некоторых антарктических рыб содержи белок, обладающий свойствами антифриза. Технология многих производств основана на переработке белков, изменении их свойств; в кожевенной промышленности, при выделке мехов, натурального шелка, выработке сыров, хлеба и т.д. Строение белковой молекулы. Для описания строения белковой молекулы были введены понятия о первичной, вторичной, третичной и четвертичной структурах белковой молекулы. В последние годы появились еще такие понятия, как сверхвторичная структура, характеризующая энергетически предпочтительные агрегаты вторичной структуры, и домены – части белковой глобулы, представляющие собой достаточно обособленные глобулярные области. Количество и последовательность расположения аминокислот, и местоположение дисульфидных связей в полипептидной цепи определяют первичную структуру белка. Между первичной структурой белка и его функцией у данного организма существует самая тесная связь. Для того, чтобы белок выполнял свойственную ему функцию, необходима совершенно определенная последовательность аминокислот в полипептидной цепи этого белка. Даже небольшие изменения в первичной структуре могут значительно изменять свойства белка и соответственно его функции. Например, в эритроцитах здоровых людей содержится белок– гемоглобин с определенной последовательностью аминокислот. Небольшая часть людей имеет врожденную аномалию структуры гемоглобина: их эритроциты содержат гемоглобин, у которого в одном положении вместо глутаминовой кислоты (заряженной, полярной) содержится аминокислота валин (гидрофобная, неполярная). Такой гемоглобин существенно отличается по физико-химическим и биологическим свойствам от нормального. Появление гидрофобной аминокислоты, приводит к возникновению «липкого» гидрофобного контакта (эритроциты плохо передвигаются в кровеносных сосудах), к изменению формы эритроцита (из двояковогнутого в серповидный), а также к ухудшению переноса кислорода и т.д. Дети, родившееся с этой аномалией, в раннем детстве погибают от серповидноклеточной анемии. Исчерпывающие доказательства в пользу утверждения, что биологическая активность определяется аминокислотной последовательностью, были получены, после искусственного синтеза фермента рибонуклеазы (Меррифилд). Синтезированный полипептид с той же аминокислотной последовательностью, что и естественный фермент, обладал такой же ферментативной активностью. Исследования последних десятилетий показали, что первичная структура закреплена генетически и в свою очередь определяет вторичную, третичную и четвертичную структуры белковой молекулы и ее общую конформацию. Первым белком, у которого была установлена первичная структура, был белковый гормон инсулин (содержит 51 аминокислоту). Это было сделано в 1953 г. Фредериком Сэнгером. К настоящему времени расшифрована первичная структура более десяти тысяч белков, но это очень небольшое количество, если учесть, что в природе белков около 1012. Зная первичную структуру белка, можно точно написать его структурную формулу, если белок представлен одной полипептидной цепью. Если в состав белка входит несколько полипептидных цепей, то их предварительно разъединяют, используя специальные реактивы. Для определения первичной структуры отдельной полипептидной цепи, методами гидролиза с использованием аминокислотных анализаторов, устанавливают ее аминокислотный состав. Затем, применяя специальные методы и реагенты, определяют природу концевых аминокислот. Для установления порядка чередования аминокислот, полипептидную цепь подвергают ферментативному гидролизу, при котором образуются осколки этой полипептидной цепи – короткие пептиды. Эти пептиды разделяют методом хроматографии и устанавливают последовательность аминокислот в каждом. Таким образом, достигается этап, когда последовательность аминокислот в отдельных пептидах (фрагментах белка) известна, но остается невыясненной последовательность самих пептидов. Последнюю устанавливают с помощью так называемых перекрывающихся пептидов. Для этого используются какой-либо другой фермент, расщепляющий исходную полипептидную цепь в других участках, и определяют аминокислотную последовательность вновь полученных пептидов. Пептиды, образованные под действием двух ферментов, содержат одинаковые фрагменты аминокислотных последовательностей., совмещая их устанавливают общую аминокислотную последовательность полипептидной цепи. Большой вклад в изучение строения белковой молекулы сделали Л.Полинг и Р.Кори. Обратив внимание на то, что в молекуле белка больше всего пептидных связей, они первыми провели кропотливые рентгеноструктурные исследования этой связи. Изучили длины связей, углы под которыми располагаются атомы, направление расположения атомов относительно связи. На основании исследований были установлены следующие основные характеристики пептидной связи.
1. Четыре атома пептидной связи и два присоединенных 2. Атомы О и Н пептидной связи и два a-углеродных атома и R-группы имеют транс-ориентацию относительно пептидной связи. 3. Длина связи С-N, равная 1,32 Å, имеет промежуточное значение между длиной двойной ковалентной связи (1,21 Å) и однородной ковалентной связи (1,47 Å). Отсюда следует, что связь С-N имеет частично характер двойной связи. Т.е. пептидная связь может существовать в виде резонансных и таутамерных структур, в кето-енольной форме.
Вращение вокруг связи –С=N– затруднено и все атомы, входящие в пептидную группу, имеют планарную транс-конфигурацию. Цис-конфигурация является энергетически менее выгодной и встречается лишь в некоторых циклических пептидах. Каждый планарный пептидный фрагмент содержит две связи с a-углеродными атомами, способными к вращению. Это связи Сa–N (угол вращения вокруг этой связи обозначается j) и связь Сa–С (угол вращения вокруг этой связи обозначается y). Пептидная связь по своей химической природе является ковалентной и придает высокую прочность первичной структуре белковой молекулы. Являясь повторяющимся элементом полипептидной цепи и имея специфические особенности структуры, пептидная связь влияет не только на форму первичной структуры, но и на высшие уровни организации полипептидной цепи. Вторичная структура белковой молекулы образуется в результате того или иного вида свободного вращения вокруг связей, соединяющих В природных полипептидных цепях обнаружены три основных типа структуры: a-спираль, складчатый лист и статистический клубок. Спиральная структура образуется если в цепи одинаковые углы поворотов (j) для всех связей Сa–N и углом поворота (y) для всех связей Сa–С и равны соответственно –48º и –57º. Наиболее часто встречается правозакрученная
Рисунок 1 – Структура и параметры a-спирали белка Спиральную структуру могут нарушить два фактора: 1) в наличие остатка пролина, циклическая структура которого вносит излом в пептидную цепь – нет группы –NН2, поэтому невозможно образования внутрицепочечной водородной связи;
2) если в полипептидной цепи подряд расположено много остатков аминокислот, имеющих положительный заряд (лизин, аргинин) или отрицательный заряд (глутаминовой, аспарагиновой кислот), в этом случае сильное взаимное отталкивание одноименнозаряженных групп (–СОО– или –NН3+) значительно превосходит стабилизирующее влияние водородных связей в Структура типа складчатого листа также стабилизирована водородными связями между теми же диполями =NН......О=С<. Однако в этом случае возникает совершенно иная структура, при которой остов полипептидной цепи вытянут таким образом, что имеет зигзагообразную структуру. Углы вращения для связей Сa-N(j) и Сa-С (y) близки соответственно к –120+1350. Складчатые участки полипептидной цепи проявляют кооперативные свойства, т.е. стремятся расположиться рядом в белковой молекуле, и формируют параллельные
одинаковонаправленные полипептидные цепи или антипараллельные,
которые укрепляются благодаря водородным связям между этими цепями. Такие структуры называются b-складчатые листы (рисунок 2).
Рисунок 2 – b-структура полипептидных цепей a-Спиральные складчатые листы – это упорядоченные структуры, в них имеется регулярная укладка аминокислотных остатков в пространстве. Участки белковой цепи с нерегулярной укладкой аминокислотных остатков в пространстве, которые также удерживаются благодаря водородным связям – называются неупорядоченными, бесструктурными – статистическим клубком. Все эти структуры возникают самопроизвольно и автоматически вследствие того, что данный полипептид имеет определенную аминокислотную последовательность, которая предопределена генетически. a-спирали и Сведения о чередовании аминокислотных остатков в полипептидной цепи, а также о наличии в белковой молекуле спирализованных, складчатых и неупорядоченных участков еще не дают полного представления ни об объеме, ни о форме, ни тем более о взаимном расположении участков полипептидной цепи по отношению друг к другу. Эти особенности строения белка выясняются при изучении его третичной структуры, под которой понимают общее расположение в пространстве в определенном объеме полипептидной цепи. Третичная структура устанавливается с помощью рентгеноструктурного анализа. Первая модель молекулы белка – миоглобина, отражающая его третичную структуру, была создана Дж. Кендрю с сотрудниками в 1957г. Несмотря на большие трудности к настоящему времени удалось установить третичную структуру более 1000 белков, в том числе гемоглобина, пепсина, лизоцима, инсулина и т.д. Третичная структура белков образуется путем дополнительного складывания пептидной цепи содержащей a-спираль, b-структуры и участки без периодической структуры. Третичная структура белка формируется совершенно автоматически, самопроизвольно и полностью предопределяется первичной структурой. Основной движущей силой в возникновении трехмерной структуры, является взаимодействие радикалов аминокислот с молекулами воды. При этом неполярные гидрофобные радикалы аминокислот группируются внутри белковой молекулы, в то время как полярные радикалы ориентируются в сторону воды. В какой-то момент возникает термодинамически наиболее выгодная стабильная конформация молекулы – глобула. В такой форме белковая молекула характеризуется минимальной свободной энергией. На конформацию возникшей глобулы оказывают влияние такие факторы как рН раствора, ионная сила раствора, а также взаимодействие белковых молекул с другими веществами. В последнее время появились доказательства, что процесс формирования третичной структуры не является автоматическим, а регулируется и контролируется специальными молекулярными механизмами. В этом процессе задействованы специфические белки – шапероны. Основными функциями их являются способность предотвращать образование из полипептидной цепи неспецифических (хаотичных) беспорядочных клубков, и обеспечение доставки (транспорта) их к субклеточным мишеням, создавая условия для завершения свертывания белковой молекулы. Стабилизация третичной структуры обеспечивается благодаря нековалентным взаимодействиям между атомными группировками боковых радикалов следующих типов: - водородные связи могут возникать между функциональными группами боковых радикалов. Например, между ОН группой тирозина и –N< в кольце остатка гистидина. - электростатические силы притяжения между радикалами, несущими противоположно заряженные ионные группы (ион-ионные взаимодействия), например отрицательно заряженная карбоксильная группа - гидрофобные взаимодействия обусловлены силами Ван-дер-Ваальса между неполярными радикалами аминокислот. (Например, группами Стабилизируется третичная структура и ковалентной дисульфидной связью (–S–S–) между остатками цистеина. Эта связь очень прочная и присутствует не во всех белках. Важную роль эта связь играет в белковых веществах зерна и муки, т.к. оказывает влияние на качество клейковины, структурно-механические свойства теста и соответственно на качество готовой продукции – хлеба и т.д. Белковая глобула не является абсолютно жесткой структурой: в известных приделах возможны обратимые перемещения частей пептидной цепи относительно друг друга с разрывом небольшого количества слабых связей и образования новых. Молекула как бы дышит, пульсирует в разных своих частях. Эти пульсации не нарушают основного плана конформации молекулы, подобно тому, как тепловые колебания атомов в кристалле не изменяют структуру кристалла, если температура не настолько велика, что наступает плавление. Только после приобретения белковой молекулой естественной, нативной третичной структуры он проявляет свою специфическую функциональную активность: каталитическую, гормональную, антигенную и т.д. Именно при образовании третичной структуры происходит формирование активных центров ферментов, центров ответственных за встраивание белка в мультиферментный комплекс, центров, ответственных за самосборку надмолекуляных структур. Поэтому любые воздействия (термические, физические, механические, химические), приводящие к разрушению этой нативной конформации белка (разрыв связей), сопровождается частичной или полной потерей белком его биологических свойств. Изучение полных химических структур некоторых белков показало, что в их третичной структуре выявляются зоны, где сконцентрированы гидрофобные радикалы аминокислот, и полипептидная цепь фактически обматывается вокруг гидрофобного ядра. Более того, в ряде случаев в белковой молекуле обособляются два и даже три гидрофобных ядра, в результате возникает 2-х или 3-х ядерная структура. Такой тип строения молекулы характерен для многих белков, обладающих каталитической функцией (рибонуклеаза, лизоцим и т.д.). Обособленная часть или область молекулы белка обладающая в определенной степени структурной и функциональной автономией называется доменом. У ряда ферментов, например, обособленны субстрат-связывающие и кофермент связывающие домены. Третичная структура белка имеет прямое отношение к его форме, которая может быть различной: от шарообразной до нитевидной. Форма белковой молекулы характеризуется таким показателем, как степень асимметрии (отношение длинной оси к короткой). У фибриллярных или нитевидных белков степень асимметрии больше 80. При степени асимметрии меньше 80 белки относятся к глобулярным. Большинство из них имеет степень асимметрии 3-5, т.е. третичная структура характеризуется достаточно плотной упаковкой полипептидной цепи, приближающейся по форме к шару. В биологическом отношении фибриллярные белки играют очень важную роль, связанную с анатомией и физиологией животных. У позвоночных на долю этих белков приходится 1/3 от их общего содержания. Примером фибрилярных белков может служить белок шелка – фиброин, который состоит из нескольких антипараллельных цепей со структурой складчатого листа. Белок a-кератин содержит от 3-7 цепей. Коллаген имеет сложную структуру, в которой 3 одинаковые левовращающие цепи скручены вместе с образованием правовращающей тройной спирали. Эта тройная спираль стабилизирована многочисленными межмолекулярными водородными связями. Наличие таких аминокислот, как гидроксипролина и гидроксилизина также вносит вклад в образование водородных связей, стабилизирующих структуру тройной спирали. Все фибриллярные белки плохо растворимы или совсем нерастворимы в воде, так как в их составе содержится много аминокислот, содержащих гидрофобные, нерастворимые в воде R-группы изолейцин, фенилаланин, валин, аланин, метионин. После специальной обработки нерастворимый и неперевариваемый коллаген превращается в желатин-растворимую смесь полипептидов, который затем используют в пищевой промышленности. Глобулярные белки выполняют разнообразные биологические функции. Они выполняют транспортную функцию, т.е. переносят питательные вещества, неорганические ионы, липиды и т.д. К этому же классу белков принадлежат гормоны, а также компоненты мембран и рибосом. Все ферменты тоже глобулярные белки.
|
||||||||||||||||||||||||
|
|
Последнее изменение этой страницы: 2017-02-05; просмотров: 330; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.222.69.152 (0.07 с.) |