
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Окислительное декарбоксилирование пирувата и цикл Кребса: последовательность реакций, связь с дыхательной цепью, регуляция, значение.
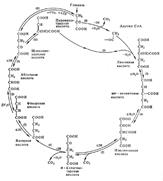
Окислительное декарбоксилирование пирувата катализируется (ПДГ). ПДГ - это комплекс из трех ферментов, для работы которого требуются следующие витамины: В1 (тиамин), В2 (рибофлавин), РР (никотинамид), пантотеновая и липоевая кислоты. ПДГ осуществляет декарбоксилирование ы) и окисление (отщепление водорода) молекулы пирувата. Продукты реакции и их дальнейшие превращения: СО2. В основном выделяется с выдыхаемым воздухом, а также используется для карбоксилирования субстратов. Ацетил-КоА. Расщепляется в цикле Кребса до СО2 и Н2; НАДН. Окисляется в дыхательной цепи. Регуляция ПДГ: ПДГ может быть в двух формах: фосфорилированной (неактивной) и дефосфорилированной (активной). АТФ, НАДН, ацетил-КоА, жирные кислоты ингибируют ПДГ, инсулин активирует. Цикл Кребса:
Таким образом, ацетил-КоА распадается в цикле Кребса до СО2 и Н2 , который присоединяется к кофакторам дегидрогеназ. В ходе цикла Кребса образуется сукцинил-КоА, который содержит макроэргическую связь (первичный макроэрг). Гидролиз этой связи сопряжен с реакцией фосфорилирования АДФ, при этом образуется АТФ. Синтез АТФ с использованием энергии макроэргической связи субстрата называется субстратным фосфорилированием. Конечные продукты цикла Кребса и пути их использования: - СО2 выдыхается с воздухом, небольшая часть используется в реакциях карбоксилирования. - НАДН и ФАДН2 окисляются в дыхательной цепи. - АТФ используется на различные виды работы: 1) механическая работа (сокращение мышц, движение сперматозоидов, лейкоцитов) 2) осмотическая работа или активный транспорт, то есть движение против градиента концентрации. 3) химическая работа, когда энергия АТФ используется в биосинтетических процессах и на активацию субстратов;4) электрическая работа (генерация биотоков); 5) при передаче гормонального сигнала (для работы аденилатциклазы и протеинкиназы). Регуляция цикла Кребса: осуществляется путем влияния на ключевые ферменты: цитратсинтазу (начинает процесс), изоцитратдегидрогеназу (лимитирующий фермент), 2-ок-соглутаратдегидрогеназу (фермент, стоящий на развилке). Цитратсинтазу активируют оксалоацетат и ацетил-КоА, ингибируют АТФ, НАДН, длинноцепочные ацилы-КоА. Изоцитратдегидрогеназа является аллостерическим ферментом. Ее активируют АДФ, Са2+, цАМФ. Ингибируют изоцитратдегидрогеназу АТФ, НАДН, НАДФН. 2-оксоглутаратдегидрогеназу активируют Са2+ и цАМФ, ингибирует сукцинил-КоА.
Значение цикла Кребса: - катаболическое и энергетическое (цикл Кребса является общим конечным путем распада для метаболитов всех классов соединений; в нем образуется АТФ в результате субстратного фосфорилирования; он является главным поставщиком водорода для дыхательной цепи); - анаболическое или биосинтетическое Промежуточные метаболиты цикла Кребса используются на синтез других соединений. Например, из оксалоацетата, 2-оксоглутарата и сукцината образуются аминокислоты; из оксалоацетата - глюкоза и другие углеводы; 11. Дыхательная цепь, компоненты, структурная организация. Электрохимический потенциал, его значение. Дыхательная цепь: Дыхательная цепь (цепь переноса электронов) - это цепь сопряженных окислительно-восстановительных реакций, в ходе которых водород, отщепленный от субстратов, переносится на кислород с образованием воды и выделением энергии. Назначение дыхательной цепи - генерирование энергии.Компоненты дыхательной цепи называются дыхательными переносчиками. Большинство из них (кроме убихинона) являются сложными белками.
Субстраты НАД-зав. Субстраты ФАД-зав. дегидрогеназ дегидрогеназ
Характеристика дыхательных переносчиков: НАДН-дегидрогеназа (НАДН-ДГ) (в схеме - ФП) - это флавинзависимый фермент, небелковой частью которого является ФМН и железо-серные центры. НАДН-ДГ встроена во внутреннюю мембрану митохондрий. Она осуществляет перенос водорода с НАДН вначале на ФМН с образованием ФМНН2, затем переносит водород с ФМНН2 на железо-серные центры и только потом на КоQ, при этом последний восстанавливается до КоQН2. Таким образом, НАДН-ДГ катализирует реакцию: НАДН2 + КоQ → НАД+ + КоQН2. КоQ (убихинон)- это небелковый переносчик, растворимый в липидах. Восстановленная форма убихинона (КоQН2) называется убихинол. Убихинон может перемещаться в липидной фазе внутренней мембраны митохондрий, представляя, таким образом, лабильный субстрат для ферментов встроенных в мембрану. Цитохромы (b, c1, c, a, а3) - это сложные белки, небелковой частью которых является гем, содержащий Fe3+. Принимая электрон, железо трехвалентное переходит в железо двухвалентное, отдавая электрон - переходит опять в трехвалентное.
12. Окислительное фосфорилирование АДФ. Механизм. Сопряжение и разобщение окисления и фосфорилирования в дыхательной цепи. Коэффициент Р/0. Регуляция дыхательной цепи. Механизм окислительного фосфорилирования. Компоненты дыхательной цепи находятся во внутренней мембране митохондрий. Они расположены таким образом, что, передавая электроны по цепи, одновременно выталкивают протоны водорода на наружную сторону мембраны в межмембранное пространство. В результате, на наружной стороне мембраны создается избыток протонов водорода (+ заряд), а с внутренней стороны – недостаток (- заряд). Это проявляется в возникновении мембранного электрохимического потенциала DmН+, который складывается из разности зарядов на мембране (Dj) и разности рН (снаружи более кислое, внутри – более щелочное). Протоны водорода могут возвращаться в матрикс по протонным каналам, с которыми связана специфическая Н+-АТФаза (АТФ-синтетаза). Обратный ток протонов по каналам в матрикс активирует этот фермент, и он катализирует синтез АТФ из АДФ и Н3РО4. Таким образом, энергия мембранного поценциала преобразуется в энергию макроэргической связи АТФ. АТФ с помощью фермента транслоказы переносится из митохондрий в цитозоль, где и используется. Сопряжение и разобщение в дыхательной цепи: Сопряжение в дыхательной цепи - это такое состояние, когда окисление (перенос электронов) сопровождается фосфорилированием, то есть синтезом АТФ. Разобщение - это такое состояние дыхательной цепи, когда окисление идет, а фосфорилирование не происходит, то есть пункты фосфорилирования выключены полностью или частично. В этом случае вся или какая-то часть образующейся энергии выделяется в виде тепла. Сопряженность дыхательной цепи можно оценить по коэффициенту Р/О. Коэффициент Р/О равен числу молей АТФ, образующихся из АДФ и Н3РО4, на 1 грамм-атом поглощенного кислорода. Разобщение в дыхательной цепи могут вызывать липофильные вещества, которые способны переносить протоны водорода с внешней стороны внутренней мембраны митохондрий на внутреннюю, минуя АТФ-синтетазу. В результате вся энергия мембранного потенциала будет рассеиваться в виде тепла. Разобщение вызывают: 2,4-ДНФ (динитрофенол), многие яды промышленных производств, бактериальные токсины. Разобщители повышают скорость переноса электронов по дыхательной цепи и выводят ее из под контроля АТФ. Регуляция дыхательной цепи:
1. АДФ стимулирует работу дыхательной цепи. Это явление называется дыхательным контролем. 2. АТФ тормозит работу дыхательной цепи и потребление кислорода. 3. Адреналин и глюкагон активируют работу дыхательной цепи. Блокаторы дыхательной цепи: 1. Ротенон блокирует дыхательную цепь на участке НАДН – КоQ. 2. Амитал, антимицин - на участке между цитохромами b и c1. 3. Цианиды и окись углерода блокируют цитохромоксидазу, при этом вся дыхательная цепь не работает. 13. Субстратное фосфорилирование АДФ. Отличия от окислительного фосфорилирования. Основные пути использования АТФ. Цикл АДФ-АТФ. Понятие о свободном окислении и его значение. Тканевые особенности окислительно-восстановительных процессов. Субстратное фосфорилирование образ-е энергии в виде АТФ за счет разрыва макроэргической связи. Отличия разные источники энергии, для окислительного необходимы движение электронов в дых цепи, для субстратного необ-ма энергия макроэргической связи. АТФ используется: 1. мех-ая работа(сокр мышц, дв-е сперматозоидов, лейкоцитов); 2 осматич-я работа или активный транспорт,т.е. движение против градиента концентрации; 3. хим работа, энергия АТФ исп-ся в биосинтет-х процессах и на активацию Субстрата; 4 электрическая (генерация биотопов); 5. при передачи гормонального сигнала (для работы аденилатциклазы и протеинкиназы). АТФ-АДФ – основной мех-м обмена энергии в биологических системах. Использование АТФ как источника энергии возможно только при условии непрерывного синтеза АТФ из АДФ за счет энергии орг соединений. Нефосфорилирующее (свободное) окисление- Это окисление без образования АТФ. Ферменты свободного окисления: оксидазы, оксигеназы, некоторые дегидрогеназы. Значение свободного окисления: - терморегуляция; - Образование биологически важных соединений (катехоламинов, глюкокортикостероидов, коллагена, активация витамина Д и т.д); - обезвреживание ксенобиотиков (ядов, токсинов, лекарств, веществ бытовой химии). Тканевые и возрастные особенности окислительных процессов. Анаэробные ткани могут получать энергию без кислорода. Такими тканями являются: скелетные мышцы, эритроциты, периферические нервы, мозговое вещество почек, кость, хрящ, соединительная ткань. Аэробные ткани получают энергию с использованием кислорода и полностью зависят от кровотока. К таким тканям относятся: головной мозг, сетчатка глаза, сердце, кора почек, печень, слизистая тонкого кишечника. Потребление кислорода, а значит, и интенсивность окислительных процессов с возрастом падают. Пути использования АТФ: 1) механическая работа (сокращение мышц, движение сперматозоидов, лейкоцитов). 2) осмотическая работа или активный транспорт (т.е. движение против градиента концентрации). 3) химическая работа, когда энергия АТФ используется в биосинтетических процессах и на актив. субстратов. 4) электрическая работа (генерация биотоков), при передаче гормонального сигнала.
|
|||||||||||||||
|
|
Последнее изменение этой страницы: 2017-01-24; просмотров: 1846; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.137.220.120 (0.012 с.) |

 Fe3+ + e- Fe2+
Fe3+ + e- Fe2+ Fe2+ - e- Fe3+
Fe2+ - e- Fe3+ Ко QН2 + 2c(Fe3+) KoQ +2Н+ + 2c(Fe2+)
Ко QН2 + 2c(Fe3+) KoQ +2Н+ + 2c(Fe2+) 2c(Fe2+) + 1/2 О2 2c(Fe3+) + О2-
2c(Fe2+) + 1/2 О2 2c(Fe3+) + О2-


