
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Синтез днк на матрице днк: пути репликации молекул, условия синтеза, его этапы.
· матрица – в ее роли выступает материнская нить ДНК, · растущая цепь – дочерняя нить ДНК, · субстраты для синтеза – dАТФ, dГТФ, dЦТФ, ТТФ, · источник энергии – dАТФ, dГТФ, dЦТФ, ТТФ, · Репликация ДНК идет в три стадии: инициация, элонгация и термина- ция. У бактерий инициация репликации ДНК начинается в уникальном сай- те хромосомы, точке репликации – oriC, из которой репликация осуществля- ется двунаправлено до точки окончания (terminus). В результате образуются две репликативные вилки, которые продвигаются в противоположных на- правлениях, т. е. обе цепи реплицируются одновременно. Праймаза синтезирует короткие РНК-праймеры для холофермента ДНК- полимеразы Ш. В месте инициации образуется промежуточный комплекс, состоящий по меньшей мере из пяти белков. Один из них – белок dnaB – может передви- гаться вдоль ДНК, используя энергию гидролиза АТР, а также служит сигна- лом для активации праймазы. Инициация репликации ДНК заканчивается образованием репликатив- ной вилки и синтезом РНК-затравки на лидирующей цепи ДНК. В процессе элонгации происходит наращивание дочерних полинуклео- тидных цепей ДНК. Каждая репликативная вилка включает, по крайней мере, две молекулы ДНК-полимеразы III, ассоциированные с несколькими вспомо- гательными белками. К последним относятся ДНК-топоизомеразы (гиразы), которые раскручивают плотно свернутую двойную спираль ДНК, и хелика- зы, которые расплетают двухтяжевую ДНК на две цепи. Ведущая цепь ДНК реплицируется непрерывно в направлении, совпа- дающем с движением репликативной вилки. Отстающая цепь считывается в направлении, противоположном движению репликативной вилки. Преодоле- ние антипараллельности цепей ДНК при репликации, возможно, достигается путем образования петельной структуры. Вначале на отстающей цепи синтезируются короткие фрагменты новой цепи ДНК, так называемые фрагменты Оказаки, названные так по имени их первооткрывателя. Каждый фрагмент начинается с короткой РНК-затравки (праймера), необходимой для функционирования ДНК-полимеразы. ДНК- полимераза III достраивает этот праймер до фрагмента ДНК длиной 1000- 2000 дезоксинуклеотидных звеньев. Кроме полимеризации цепей, которую осуществляет Рol III, в ходе элонгации ДНК происходят следующие события: 1) вырезание РНК-праймеров из лидирующей цепи и из каждого фраг- мента Оказаки. Эту функцию выполняет Pol I, благодаря 5→3- экзонуклеазной активности; 2) заполнение брешей, оставшихся после удаления праймеров. В этом процессе участвует также ДНК-полимераза I, используя для встраивания нуклеотидов 3-ОН-группу соседнего фрагмента Оказаки (рис. 29.8); 3) соединение фрагментов ДНК в отстающей цепи с помощью фермен- та ДНК-лигазы; 4) исправление ошибок репликации, благодаря 3→5- экзонуклеазной активности, которой обладают как Pol III, так и Pol I. Терминация синтеза ДНК наступает вследствие исчерпания матрицы. Репликационные «глазки» сливаются, и на каждой матрице образуется до- черняя цепь ДНК.
|
|||||
|
|
Последнее изменение этой страницы: 2017-01-24; просмотров: 447; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.222.163.31 (0.003 с.) |
 Каждая из цепей двойной спирали ДНК служит матрицей для репликации комплементарных дочерних цепей. При этом образуются две дочерние двухцепочечные молекулы ДНК, идентичные родительской, причем каждая из этих молекул содержит одну неизменную цепь родительской ДНК. Этот механизм репликации ДНК, на- званный полуконсервативным. Как любой матричный биосинтез, репликация требует наличия нескольких компонентов:
Каждая из цепей двойной спирали ДНК служит матрицей для репликации комплементарных дочерних цепей. При этом образуются две дочерние двухцепочечные молекулы ДНК, идентичные родительской, причем каждая из этих молекул содержит одну неизменную цепь родительской ДНК. Этот механизм репликации ДНК, на- званный полуконсервативным. Как любой матричный биосинтез, репликация требует наличия нескольких компонентов: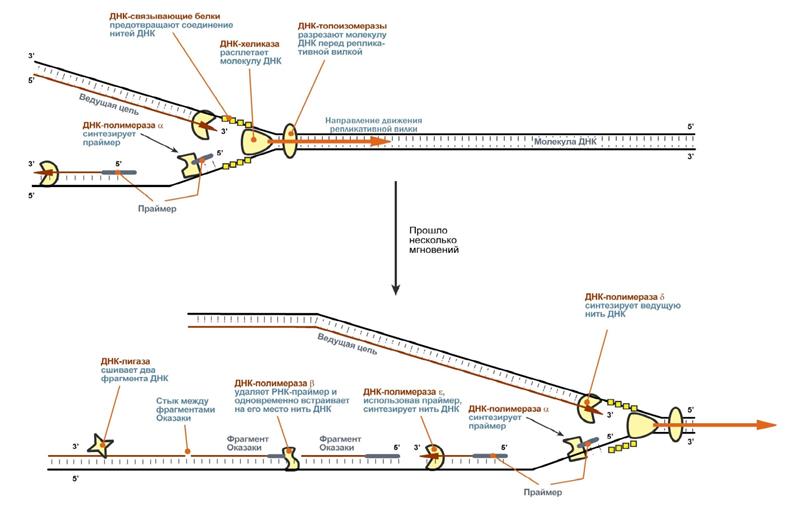 ферменты.
ферменты.


