
Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Тема 9. 2. Переваривание белков в желудке и кишечнике, всасывание аминокислот
1. При переваривании происходит гидролиз пищевых белков до свободных аминокислот. Процесс переваривания начинается в желудке и продолжается в тонком кишечнике под действием ферментов пептидгидролаз (пептидаз). Основные пептидазы синтезируются в клетках желудка, поджелудочной железы и кишечника (рис. 9.1). В желудке белки пищи денатурируются и гидролизуются с образованием олигопептидов. В кишечнике панкреатические пептидазы продолжают гидролиз олигопептидов до ди- и трипептидов и свободных аминокислот. Короткие пептиды расщепляются до свободных аминокислот в пристеночном слое или в клетках кишечного эпителия. Затем происходит их всасывание. Все пептидазы, в зависимости от места расположения в пептиде гидролизуемой связи, делятся н а эндопептидазы и экзопептидазы: • эндопептидазы - расщепляют пептидные связи, удаленные от концов пептидной цепи (пепсин, трипсин, химотрипсин, эластаза); • экзопептидазы -гидролизуют пептидные связи, образованные N- и С-концевыми аминокислотами (аминопептидаза, карбоксипептидазы А и В), а также расщепляют ди- и трипептиды.
Ферменты, участвующие в переваривании белков, обладают относительной субстратной специфичностью и гидролизуют пептидные связи; при этом каждая пептидаза преимущественно расщепляет связи, образованные определенными аминокислотами (табл. 9.1). 2. Желудочные и панкреатические пептидазы вырабатываются в неактивной форме (проферменты), секретируются к месту действия, где активируются путем частичного протеолиза (отщепление пептида различной длины, чаще с N-конца молекулы профермента). Место синтеза проферментов (клетки слизистой оболочки желудка, поджелудочная железа) и место их активации (полость желудка, тонкого кишечника) пространственно разделены. Такой механизм образования активных ферментов необходим для защиты секреторных клеток желудка и поджелудочной железы от самопереваривания. Таблица 9.1. Характеристика протеолитических ферментов желудочно-кишечного тракта
Преждевременная активация проферментов в секреторных клетках происходит при: • язвенной болезни желудка - пепсиноген превращается в пепсин в клетках слизистой желудка;
• остром панкреатите - трипсиноген превращается в трипсин в клетках поджелудочной железы и активирует остальные панкреатические пептидазы. 3. Слизистая оболочка желудка вырабатывает следующие факторы, необходимые для переваривания белков: • пепсиноген - синтезируется в главных клетках; • соляную кислоту - вырабатывается обкладочными клетками. Гидролиз белков в желудке происходит под действием пепсина. Профермент пепсиноген при поступлении пищи секретируется в полость желудка, где в два этапа происходит его активация: • с помощью НС! - медленно; • аутокаталитически - быстро, уже имеющимся пепсином. Соляная кислота желудочного сока выполняет следующие функции: •.участвует в активации пепсиногена; • создает оптимум рН для пепсина; • оказывает бактерицидное действие; • денатурирует белки пищи. Значение рН желудочного сока в норме составляет 1,5-2,0. Определение кислотности желудочного сока используется для диагностики различных заболеваний желудка (табл. 9.2). Таблица 9.2. Компоненты желудочного сока в норме и при патологических состояниях • • Связанная соляная кислота - это соляная кислота, связанная с белками и продуктами их переваривания. • Свободная соляная кислота - это соляная кислота, не связанная с другими молекулами. • Общая кислотность желудочного сока выражается в титрационных единицах (ТЕ) и измеряется количеством 0,1 М NaOH в миллилитрах, затраченным на титрование 100 мл желудочного сока в присутствии определенных индикаторов. • Кислотность желудочного сока в норме составляет: общая 40-60 ТЕ; связанная HCl 20-30 ТЕ; свободная HCl 20-40 ТЕ. При диагностике заболеваний желудка помимо биохимических анализов обязательно проводят рентгенологические и эндоскопические исследования, иногда биопсию. В слизистой оболочке желудка вырабатывается также внутренний фактор (фактор Касла), который представляет собой белок, способствующий всасыванию витамина В12 в тонкой кишке. Отсутствие этого витамина часто приводит к развитию анемии.
Молочная кислота в норме в желудочном соке отсутствует. Образуется при уменьшении содержания или отсутствии свободной соляной кислоты в результате усиленного размножения молочнокислых бактерий или при злокачественных опухолях желудка. НС1 и пепсин способны разрушать клетки эпителия желудка. В норме это не происходит благодаря наличию защитных факторов слизистой оболочки, таких, как: • образование на поверхности слизи, содержащей гетерополисахариды, которые не являются субстратами пептидгидролаз; • секреция эпителиальными клетками ионов НСО3-, создающих в пристеночном слое менее агрессивную среду с рН 5,0-6,0, в которой пепсин неактивен. Кроме того, клетки поврежденного эпителия обладают способностью к быстрой регенерации. Пепсин гидролизует пептидные связи, образованные аминогруппой или карбоксильной группой ароматических аминокислот (см. табл. 9.1):
• ферментов поджелудочной железы-трипсина, химотрипсина, эластазы, карбоксипептидаз А и Б; • ферментов эпителия тонкой кишки - аминопептидазы, дипептидаз, трипептидаз. Активная форма трипсина образуется в кишечнике при участии фермента энтеропептидазы, выделяемого клетками кишечника. Энтеропептидаза отщепляет от N-конца трипсиногена гексапептид (рис. 9.2), что приводит к изменению конформации молекулы и формированию активного центра трипсина.
Пунктирная стрелка - место гидролиза. Буквами обозначены аминокислоты (А - асп, Г - глу, В - вал, Л - лиз, И - иле) Остальные проферменты панкреатического сока (химотрипсиноген, прокарбоксипептидазы А и В, проэластаза) активируются трипсином. Активация панкреатических пептидаз в кишечнике происходит в виде каскада реакций.
• Ферменты, участвующие в переваривании белков в кишечнике, обладают субстратной специфичностью к определенным аминокислотам в белке: Трипсин гидролизует преимущественно пептидные связи, образованные карбоксильными группами катионогенных аминокислот: • • • Карбоксипептидазы отщепляют С-концевые аминокислоты: карбоксипептидаза А - гидрофобные аминокислоты, карбоксипептидаза В - Лиз и Арг; • Аминопептидаза - отщепляет N-концевые аминокислоты; • Дипептидаза - гидролизует дипептиды; • Трипептидаза - расщепляет трипептиды на отдельные аминокислоты. 5. Конечным результатом переваривания белков является образование свободных аминокислот, поступающих в клетки слизистой оболочки кишечника, путем вторично-активного транспорта за счет градиента концентрации натрия (симпорт). Всасывание L-аминокислот является активным, требующим затраты энергии процессом. Транспорт их осуществляется двумя путями: • через воротную систему печени; • по лимфатическим сосудам, сообщающимся с кровью через грудной лимфатический проток.
Аминокислоты конкурируют друг с другом за специфические участки связывания белков-переносчиков. Так, всасывание лейцина в больших количествах уменьшает всасывание изолейцина и валина. В крови максимальная концентрация аминокислот достигается через 30-50 минут после приема белковой пищи. Свободные аминокислоты, в отличие от белков пищи, лишены видовой специфичности и не обладают антигенными свойствами. Скорость проникновения аминокислот через мембраны клеток различается, что указывает на существование транспортных систем, обеспечивающих перенос аминокислот через мембраны. Известно пять специфических транспортных систем для переноса определенной группы близких по строению аминокислот: • нейтральных с короткой боковой цепью (аланин, серин, треонин); • нейтральных с длинной или разветвленной боковой цепью (валин, лейцин, изолейцин); • с катионными радикалами (лизин, аргинин); • с анионными радикалами (глутаминовая и аспарагиновая кислоты); • иминокислот (пролин, оксипролин). Переносчики аминокислот первой и пятой групп, а также переносчик метионина относятся к числу Na+-зависимых. Независимые от Na+ переносчики специфичны для некоторых нейтральных аминокислот (фенилаланин, лейцин) и аминокислот с катионными радикалами (лизин). Одной из основных транспортных систем для нейтральных аминокислот является γ-глутамильный цикл, который функционирует в почках, поджелудочной железе, печени и селезенке, в мозге и других тканях он содержится в очень небольших количествах (рис. 9.3).
Е1 - γ-глутамилтранспептидаза (γ-ГТ); Е2 - γ-глутамилциклотрансфераза; Е3 - пептидаза; Е4 - оксопролиназа; Е5 - γ-глутамилцистеинсинтетаза; Е6 - глутатионсинтетаза. Система состоит из одного мембранного и пяти цитоплазматических ферментов. Перенос аминокислоты внутрь клетки осуществляется в комплексе с глутамильным остатком глутатиона под действием фермента γ-ГТ. Затем аминокислота освобождается, а γ-глутамильный остаток в несколько стадий превращается в глутатион, который способен присоединять следующую молекулу аминокислоты Ключевую роль в работе системы играет мембранный фермент γ -глутамилтранспептидаза (γ-ГТ). Этот гликопротеин катализирует перенос γ-глутамильной группы с глутатиона (γ -глутамилцистеинилглицин) или другого γ-глутамильного пептида на транспортируемую аминокислоту и последующий перенос комплекса в клетку. Глутатион содержится во всех тканях животных. Для транспорта в клетку одной молекулы аминокислоты с участием γ-глутамильного цикла затрачивается три молекулы АТР.
γ-ГТ в больших количествах содержится в почках, поджелудочной железе, печени и других тканях. Активность фермента в сыворотке крови в норме составляет 30-50 МЕ/л (мкмоль/мин-мг) для мужчин и 25-35 МЕ/л для женщин. Определение активности γ-ГТ в сыворотке крови используется для диагностики заболеваний печени и сердца. γ-ГТ-тест используется также в качестве маркера рака поджелудочной железы, печени, предстательной железы и для обнаружения людей из группы раннего риска алкоголизма, для контроля за лечением хронического алкоголизма.
|
|||||||||||||
|
|
Последнее изменение этой страницы: 2017-01-24; просмотров: 1159; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 18.118.145.114 (0.017 с.) |
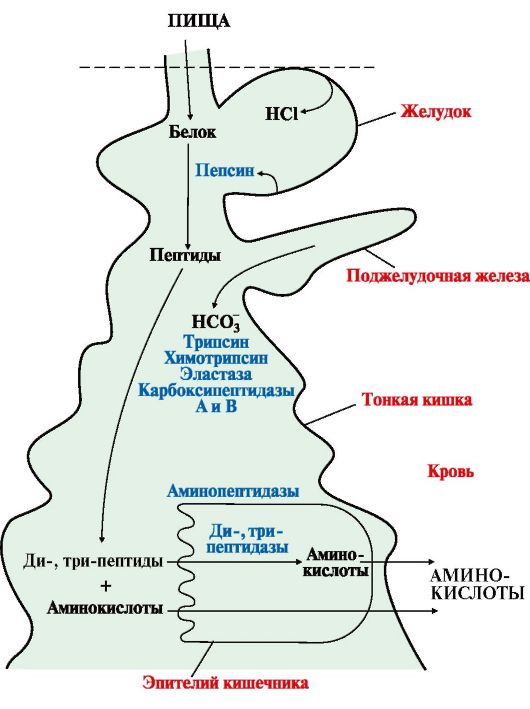 Рис. 9.1. Переваривание белков в желудочно-кишечном тракте
Рис. 9.1. Переваривание белков в желудочно-кишечном тракте Примечание: X - любая аминокислота
Примечание: X - любая аминокислота Общая кислотность желудочного сока - это совокупность всех кислотореагирующих веществ желудочного сока, представляющая собой секрет желудка, собираемый в течение 1 часа (предварительно отбирают секрет, ранее содержащийся в желудке).
Общая кислотность желудочного сока - это совокупность всех кислотореагирующих веществ желудочного сока, представляющая собой секрет желудка, собираемый в течение 1 часа (предварительно отбирают секрет, ранее содержащийся в желудке). 4. Переваривание белков в кишечнике происходит под действием:
4. Переваривание белков в кишечнике происходит под действием: Рис. 9.2. Механизм активации трипсиногена.
Рис. 9.2. Механизм активации трипсиногена. Ферменты эпителия тонкой кишки синтезируются в энтероцитах сразу в активной форме.
Ферменты эпителия тонкой кишки синтезируются в энтероцитах сразу в активной форме. Химотрипсин - пептидные связи, образованные карбоксильными группами ароматических аминокислот:
Химотрипсин - пептидные связи, образованные карбоксильными группами ароматических аминокислот: Эластаза - связь между -Гли-Ала-
Эластаза - связь между -Гли-Ала-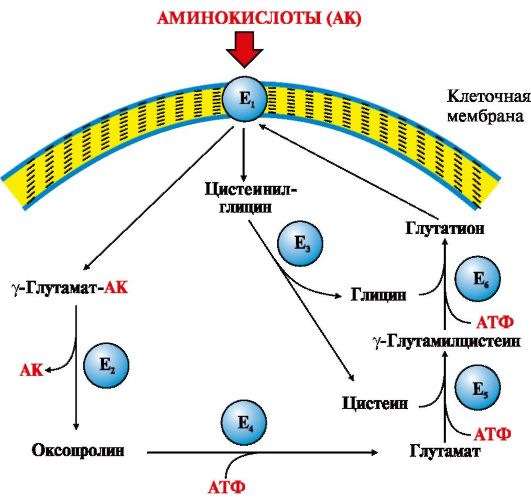 Рис. 9.3. γ-Глутамильный цикл:
Рис. 9.3. γ-Глутамильный цикл:


