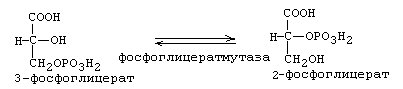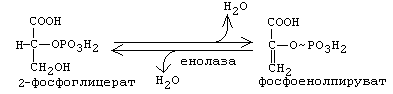Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Механизмы трансмембранного переноса глюкозы
Транспорт моносахаридов из просвета кишечника в клетки слизистой оболочки может осуществляться путем облегченной диффузии и активного транспорта. При активном транспорте глюкоза и Na+ проходят с люминальной стороны, связываясь с разными участками белка-переносчика. При этом Na+ поступает в клетку под влиянием электрохимического градиента и «тащит» глюкозу за собой. Следовательно, чем больше градиент Na+, тем больше поступление глюкозы. Если концентрация Na+ во внеклеточной жидкости уменьшается, транспорт глюкозы подавляется. Градиент концентрации Na+ являющийся движущей силой этого симпорта, создается работой Na+K+ нacoca. Глюкоза из клетки кишечника затем перемещается во внеклеточную жидкость и далее в кровь с помощью облегченной диффузии. Поступающая из кишечника глюкоза с кровью воротной вены попадает в печень, где часть ее задерживается, а часть через общий кровоток попадает в клетки других органов и тканей. Потребление глюкозы клетками из кровотока происходит также путем облегченной диффузии при участии специальных белков-транспортеров. Следовательно, скорость трансмембранного потока глюкозы зависит только от градиента ее концентрации. Исключением являются клетки мышц и жировой ткани, где облегченная диффузия регулируется инсулином. В отсутствие инсулина плазматическая мембрана этих клеток непроницаема для глюкозы. Белки-переносчики (транспортеры глюкозы — ГЛЮТ) обнаружены во всех тканях. Существует несколько разновидностей ГЛЮТ, которые пронумерованы по порядку их обнаружения. Все 5 типов ГЛЮТ имеют сходную первичную структуру. ГЛЮТ-1 служит для обеспечения стабильного потока глюкозы в мозг. В других тканях он поставляет глюкозу в клетки, когда они находятся в стоянии покоя. ГЛЮТ-2 обнаружен в клетках органов, выделяющих глюкозу в кровь. Именно при участии ГЛЮТ-2 глюкоза переходит в кровь из энтероцитов после всасывания в кишечнике. ГЛЮТ-3 обладает большим, чем ГЛЮТ-1, сродством к глюкозе. Он также обеспечивает постоянный приток глюкозы к клеткам нервной ткани. ГЛЮТ-4 — главный переносчик глюкозы в мышцах и адипоцитах. ГЛЮТ-5 встречается главным образом в клоках тонкой кишки. Его функции известны недостаточно. Все типы ГЛЮТ могут находиться как в плазматической мембране, так и в цитозольных везикулах.
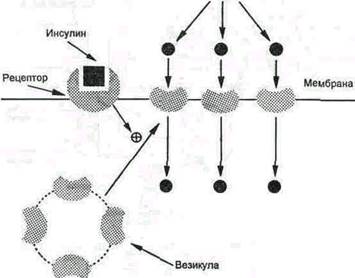
В отсутствие инсулина ГЛЮТ-4 (и в меньшей мере ГЛЮТ-1) почти полностью находится в цитоплазме (рис.3). Влияние инсулина на такие клетки приводит к перемещению везикул, содержащих ГЛЮТ, к плазматической мембране и их слиянию с ней, после чего возможен облегченный транспорт глюкозы в эти клетки. После снижения концентрации инсулина в крови белки-транспортеры глюкозы снова перемещаются в цитозоль. В клетки печени глюкоза проходит при участии ГЛЮТ-2, независимого от инсулина. Концентрация глюкозы в гепатоцитах в период пищеварения повышается соответственно ее уровню в крови воротной вены. Хотя инсулин и не влияет на транспорт глюкозы, он усиливает приток глюкозы в гепатоцит в период пищеварения косвенным путем, индуцируя синтез глюкокиназы и ускоряя тем самым фосфорилирование глюкозы.
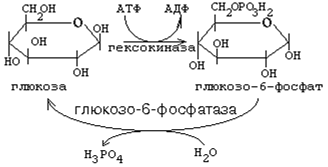
Рисунок 3 Транспорт глюкозы из первичной мочи в клетки канальцев происходит путем активного транспорта подобно тому, как это происходит с люминальной стороны кишечника в клетки. Благодаря этому глюкоза может поступать в клетки даже в том случае, если ее концентрация в просвете кишечника или в первичной моче меньше, чем в клетках. Глюкоза реабсорбируется из первичной мочи почти полностью (на 99%) к конечной части канальцев. Метаболизм глюкозы в клетке Первая реакция, в которую вступает глюкоза в клетке, является единственной. Это реакция фосфорилирования глюкозы за счёт АТФ. Фермент, катализирующий эту реакцию, есть в любой клетке. Он называется гексокиназа (ГК).
Биологический смысл гексокиназной реакции: 1. Сделать молекулу глюкозы более способной к химическим реакциям, ослабить в ней химические связи, дестабилизировать её ("расшатать"). 2. Связать, задержать глюкозу в клетке, чтобы она не смогла выйти обратно в кровь (глюкозо-6-фосфат не способен проходить через клеточную мембрану). Чтобы связанная молекула могла выйти из клетки, глюкозо-6-фосфат должен превратиться обратно в глюкозу. Фермент, катализирующий обратную реакцию (превращение глюкозо-6-фосфата обратно в глюкозу), называется глюкозо-6-фосфатаза. Он гидролизует глюкозо-6-фосфат до глюкозы и Н3РО4 (Фн), то есть катализирует обходной обратный путь гексокиназной реакции. Глюкозо-6-фосфатаза есть в печени, почках и слизистой оболочке кишечника.
3. Гексокиназа - это ключевой фермент всего метаболизма глюкозы. Он лимитирует (ограничивает) скорость всех путей метаболизма глюкозы в клетке. Гексокиназа всегда работает с максимальной скоростью. Только в печени есть ещё один фермент, катализирующий реакцию превращения глюкозы в глюкозо-6-фосфат. Это изофермент гексокиназы - глюкокиназа. "На высоте пищеварения", когда концентрация глюкозы в воротной вене во много раз возрастает, глюкокиназа работает очень интенсивно. Следовательно, утилизация глюкозы клетками печени "на высоте пищеварения" возрастает при подключении дополнительного пути метаболизма (работа глюкокиназы). Регуляторная роль гексокиназы: этот фермент угнетается избытком своего продукта - глюкозо-6-фосфата. Если по какой-то причине дальнейшее использование глюкозо-6-фосфата замедляется (его концентрация при этом возрастает), то автоматически тормозится гексокиназная реакция. Поэтому в такой ситуации замедляется использование в клетке глюкозы в целом. После образования глюкозо-6-фосфата начинается разветвление дальнейших путей метаболизма глюкозы. Таких главных путей три. 1. Гексозомонофосфатный путь распада углеводов (ГМФ-путь) 2. Гексозобисфосфатный путь распада углеводов (ГБФ-путь). 3. Синтез гликогена. Есть ещё минорные пути (в них используется небольшая доля глюкозы, поступающей в клетку). Эти пути не играют энергетической роли, а используются для построения олиго- и полисахаридных цепей гликопротеинов, то есть выполняют структурную роль.
Г Б Ф - П У Т Ь. (ГЕКСОЗОБИСФОСФАТНЫЙ ПУТЬ РАСПАДА УГЛЕВОДОВ)
БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ГБФ-ПУТИ. 1. Это главный путь распада углеводов до конечных продуктов. Во многих клетках это - единственный путь. Так распадается 70-75% глюкозы, которая поступает в клетку. 2. Только ГБФ-путь дает клетке энергию в виде АТФ. Это основной источник получения энергии в клетке. 3. Это самый длинный путь распада углеводов. ГБФ-путь делим на 3 этапа. 1-й этап протекает в цитоплазме, дает 8 молекул АТФ при распаде 1 молекулы глюкозы или 9АТФ при распаде одного глюкозного фрагмента гликогена. Заканчивается образованием 2-х молекул пирувата (ПВК). 2-й и 3-й этапы - (исключительно аэробные!) в митохондриях с обязательным участием кислорода. 2-й этап ГБФ-пути называется "окислительное декарбоксилирование пирувата". Пируват превращается в ацетил-коэнзим А. Молекула ацетил-коэнзима А вступает в 3-й этап. Этот 3-й этап называется циклом трикарбоновых кислот (ЦТК). В этом цикле АцКоА полностью расщепляется до СО2 и Н2О. 2-й и 3-й этапы дают 30 АТФ в расчете на одну молекулу глюкозы. 1-й этап проходит 10 промежуточных стадий. В ходе первой части этого этапа молекула глюкозы расщепляется пополам до 2-х молекул фосфоглицеринового альдегида (ФГА). ОСОБЕННОСТИ ПЕРВОЙ ЧАСТИ 1-ГО ЭТАПА: Гексокиназа работает, чтобы ослабить прочную молекулу глюкозы:
2-я реакция - изомеризации:
На 3-й стадии фруктозо-6-фосфат еще более ослабляется фосфофруктокиназой (ФФК) и образуется фруктозо-1,6-бисфосфат:
Фосфофруктокиназа - это ключевой фермент ГБФ-пути. Он является "пунктом вторичного контроля". Избыток АТФ и избыток цитрата сильно ингибируют ФФК. Из-за угнетения ФФК накапливаются глюкозо-6-фосфат (Г-6-Ф) и фруктозо-6-фосфат(Ф-6-Ф). глюкозо-6-фосфат ингибирует гексокиназу, уменьшая утилизацию глюкозы клеткой и одновременно активирует гликогенсинтетазу. Если нет избытка АТФ и цитрата, а есть избыток АДФ, то АДФ активирует ФФК, и тогда скорость всего ГДФ-пути лимитируется опять гексокиназой. В результате фосфофруктокиназной реакции молекула фруктозо-1,6-бисфосфата распадается на 2 триозы при участии фермента альдолазы (4-я реакция):
5-я реакция:
В следующую (шестую) реакцию ГБФ-пути вступает только ФГА. В результате уменьшается его концентрация и равновесие 5-й реакции сдвигается в сторону образования ФГА. Постепенно весь ФДА переходит в ФГА, и поэтому количество АТФ, синтезировавшееся в последующих реакциях ГБФ-пути, мы учитываем в расчете на 2 молекулы ФГА и других промежуточных метаболитов, которые из него образуются. В 1-й части 1-ого этапа (от глюкозы до ФГА) расходуется 2 молекулы АТФ: одна - в гексокиназной реакции, другая - в фосфофруктокиназной (3-я реакция первого этапа ГБФ-пути). 2-я часть первого 1-го этапа начинается с окисления ФГА до ФГК (фосфоглицериновой кислоты) в 6-й реакции.
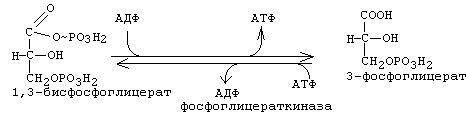
Эта реакция катализируется ферментом "глицеральдегидфосфатдегидрогеназа". Отщепляемый водород передается на НАД с образованием НАДН2. Энергии, которая выделяется при этом окислении, хватает и на то, чтобы одновременно обеспечить присоединение фосфата к альдегидной группе. Присоединяется фосфат макроэргической связью. В результате образуется 1,3-дифосфоглицериновая кислота (1,3-бисфосфоглицерат). 7-я реакция: субстратное фосфорилирование.
Фосфат с макроэргической связью передается на АДФ с образованием АТФ. В результате 7-й стадии в молекуле фосфоглицериновой кислоты остается 1 остаток фосфорной кислоты. 8-я реакция: Фосфат переносится из 3-го во второе положение и образуется 2-фосфоглицериновая кислота.
9-я реакция:
От 2-фосфоглицериновой кислоты отнимается Н2О. Это приводит к перераспределению молекулярной энергии. В результате на фосфате во втором положении накапливается энергия и связь становится макроэргической. Получается фосфоенолпируват(ФЕП).
10-я реакция: Субстратное фосфорилирование. Фосфат переносится на АДФ с образованием АТФ. ФЕП переходит в ПВК (пировиноградную кислоту).
На этом 1-й этап ГДФ-пути заканчивается, ПВК уходит в митохондрию и вступает во второй этап ГДФ-пути. Итоги 1-го этапа: 10 реакций, из которых первая, третья и десятая реакции необратимы. Сначала расходуется 2 АТФ на 1 молекулу глюкозы. Потом окисляется фосфоглицероальдегид. Энергия реализуется в ходе 2-х реакций субстратного фосфорилирования: в каждой из них образуется по 2 АТФ. Следовательно, на каждую молекулу глюкозы (на 2 молекулы ФГА) получается 4 АТФ путем субстратного фосфорилирования. Суммарно все 10 стадий можно описать следующем уравнением: С6Н12О6 + 2Н3РО4 + 2АДФ + 2НАД -----> 2С3Н4О3 + 2АТФ + 2Н2О + 2НАДН2 НАДН2 по системе митохондриального окисления передает водород на кислород воздуха с образованием Н2О и 3 АТФ, но 1-й этап протекает в цитоплазме и НАДН2 не может проходить через мембрану митохондрий. Существуют челночные механизмы, обеспечивающие этот переход НАДН2 через митохондриальную мембрану - малат-аспартатный челнок и глицерофосфатный челнок В расчете на одну молекулу глюкозы образуется 2НАДН2.
В дополнение к 2 АТФ, получаемым на 1-м этапе путем субстратного фосфорилирования, образуется еще 6 АТФ с участием кислорода, итого - 8 молекул АТФ. Столько АТФ образуется в расчете на каждую расщепленную до ПВК молекулу глюкозы в ходе первого этапа ГБФ-пути. Если эти 8 АТФ добавить к 30 молекулам АТФ, которые образуются на 2-м и 3-м этапах, то суммарный энергетический итог всего ГБФ-пути составит 38 АТФ на каждую молекулу глюкозы, расщепленную до СО2 и Н2О. В этих 38 АТФ заключено 65 процентов энергии, которая выделилась бы при сжигании глюкозы на воздухе. Это доказывает очень высокую эффективность работы ГБФ-пути. Из 38 АТФ основная их часть образуется на 2-м и 3-м этапах. Каждый из этих этапов абсолютно необратим и требует обязательного участия кислорода, так как окислительные стадии этих этапов сопряжены с митохондриальным окислением (без него невозможны). Весь ГБФ-путь от глюкозы или гликогена до СО2 и Н2О называют: АЭРОБНЫЙ РАСПАД УГЛЕВОДОВ. Ключевые ферменты первого этапа ГБФ-пути: ГЕКСОКИНАЗА и ФОСФОФРУКТОКИНАЗА.
|
|||||||||||||
|
|
Последнее изменение этой страницы: 2017-01-19; просмотров: 191; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.137.185.180 (0.032 с.) |