Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Полное и неполное окисление субстрата.
При аэробном дыхании конечным акцептором электронов в цепи переноса является молекулярный кислород, поэтому для облигатно аэробных микроорганизмы он — необходимое ростовое вещество. Молекулярный кислород необходим: • в реакциях включения одного-двух атомов кислорода в некоторые молекулы (например, стероидов, отдельных жирных кислот, каротиноидов; • для начальных этапов окисления некоторых субстратов (например, метана у метилотрофов и ароматических молекул при их расщеплении) с помощью моно- и диоксигеназ. Следует заметить, что сами эти реакции не дают энергии, но они помогают дальнейшему окислению субстрата; • при работе оксидаз (например, глюкрзооксидазы, цитохромоксидазы); • как конечный акцептор электронов в дыхательной цепи. Окончательное окисление может произойти, если тот или иной субстрат сначала превратится в центральный интермедиат ацетил-КоА или другие интермедиаты ЦТК. Для окисления белков микроорганизмы выделяют внеклеточные протеазы, которые гидролизуют белки до коротких пептидов и аминокислот.
Краткая характеристика важнейших микроорганизмов. участвующих в аэробном окислении белков (аммонификация), углеводов, углеводородов и других многоуглеродных веществ. Разложение белка микроорганизмами (аммонификации) всегда сопровождается образованием ряда продуктов: аммиака при дезаминировании аминокислот, сероводорода при расшеплении серусодержащих аминокислот (метионина, цистеина и их производных) и индола при распаде триптофана сопровождается подщелачиванием среды. Аминокислоты дезаминируются с образованием органических кислот (пирувата, ацетата и других интермедиатов ЦТК) и в таком виде входят в цикл Крэбса для полного окисления и получения клеткой энергии. Также они могут быть использованы как источник углерода для синтезов клеточных компонентов. Аминогруппы аминокислот идут либо на анаболизм азотсодержащих веществ клетки, либо выделяются в виде иона аммонии. Липиды расщепляются микробными липазами до глицерола и жирных кислот. Глицсрол затем фосфорилируется, окисляется до ДОАФ и катаболизируется через реакции гликолиза. Жирные кислоты подвергаются окислению до ацетил-КоА.
Целлюлоза гидролизуется с помощью внеклеточных целлюлаз (в аэробных условиях — грибами и миксобактериями, в анаэробных — клостридиями и некоторыми анаэробными грибами).
Ксилан древесины и луба гидролизуется ксиланазами до ксилозы+арабинозы. Есть соответствующие ферменты для гидролиза крахмала, пектина, фруктозанов, маннанов, агара, хитина, лигнина. Дисахариды далее подвергаются гидролизу или фосфотолизу. Моносахара превращаютмя в пируват гликолизм, ПФП или КДФГ. Липиды расщепляются до глицерола и жирных кислот липазами. Глицерол затем фосфорилируется. Жирные кислоты подвергаются окислению до ацетил-КоА. Ароматические соединения сначала расщепляются моно- или ди-оксигеназами до (i-кетоадипиновой кислоты или ее производных, которые затем разлагаются на ацетил-КоА и сукцинат. Предельные и непредельные углеводороды используются почвенными микроорганизмами разных групп, которые сначала окисляют их до соответствующих жирных кислот с помощью оксигеназ а затем подвергают р-окислению (кроме одноуглеродных соединений).
Неполное окисление – конечный продукт не CO2. Конечные продукты неполного окисления – орг соединения(ацетат). Неполное окисление – у уксуснокислых бактерий. Acetobacter, Gluconobacter.
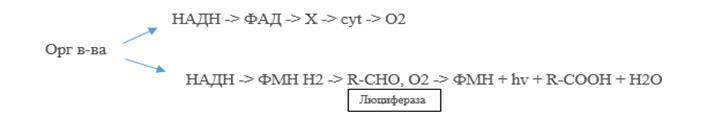
Светящиеся бактерии. Одна из групп бактерий, использующих многоуглеродные субстраты, отличается уникальной способностью к биолюминесценции. Это светящиеся бактерии — морские организмы, хемоорганотрофы. Они грамотрицательны, факультативно анаэробные палочки, которые передвигаются с помощью жгутиков. Свечение наблюдается только в присутствии кислорода. Обычно бактерии испускают сине- зеленый свет (около 500 нм), но один штамм Vibrio fischeri светится желтым светом (545 нм). В аэробных условиях микроорганизмы осуществляют процесс аэробного дыханияи свечения. У них имеется обычная дыхательная цепь и работает цикл Кребса. Свечение зависит от окисления длинноцепочечного альдегида с 13-18 атомами углерода в молекуле. Светится возбужденный флавин под действием фермента люциферазы. Люцифераза — это двухсубъединичный фермент типа монооксигеназы. Биологический смысл свечения пока непонятен. Есть гипотеза, что это защита от переокисления: когда накапливается много восстановленного пиридиннуклеотида, срабатывает «отводной канал», электроны сбрасываются на флавин и высвечиваются. В настоящее время показано, что способностью к свечению обладают представители родов Photobacterium, Beneckia и Vibrio.
Метилотрофы. Метилотрофные микроорганизмы — физиологическая группа, способная расти на одноуглеродных соединениях. Одноуглеродными называют такие соединения углерода, в состав молекулы которых входит один атом углерода или их может быть несколько, но при этом молекула никогда не содержит С-С-связей. Наибольшее значение имеют такие С1-соединения как метан, выделяемый метаногенами, метанол, образуемый при разложении пектина и лигнина, продукт смешанного брожения формиат и метиламины, получающиеся при гибели морских животных и растений. Формальдегид в свободном виде не встречается. Формиат никогда не накапливается в больших количествах. Триметиламин образуется из N- окиси триметиламина, которая синтезируется у морских обитателей как осморегулятор, поддерживающий в цитоплазме их клеток осмотическое равновесие. В анаэробных условиях С1-соединения используются различными группами микроорганизмов. Метан в отсутствии кислорода может окисляться до СО2 только ассоциацией по крайней мере двух микроорганизмов – метаногенного архея и сульфатредуцирующей бактерии, и анаэробное окисление метана количественно связано с восстановлением сульфата.
Метанол в анаэробной зоне используется сульфатредукторами, гомоацетогенами или метаногенами. Формиат может расщепляться самими энтеробактериями, образующими его при смешанном брожении. Окисление метана в аэробных условиях происходит в соответствие со схемой: СН4 → СН3ОН → НСОН → НСООН → СО2. Метилотрофы подразделяют на факультативных и облигатных. Факультативные метилотрофы способны использовать как одно-, так и многоуглеродные соединения. Облигатные метилотрофы растут только на С1-соединениях. Среди них выделяют группу, способную окислять только метан (метанотрофы). Другие одноуглеродные соединения (обычно метанол) они окисляют значительно реже и менее эффективно. В природе метилотрофы играют важную роль в глобальном цикле углерода. Они являются бактериальным газовым фильтром для метана, выделяемого в атмосферу метаногенными археями.
|
||||||
|
|
Последнее изменение этой страницы: 2016-12-30; просмотров: 1414; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.135.195.249 (0.005 с.) |