Заглавная страница Избранные статьи Случайная статья Познавательные статьи Новые добавления Обратная связь КАТЕГОРИИ: ТОП 10 на сайте Приготовление дезинфицирующих растворов различной концентрацииТехника нижней прямой подачи мяча. Франко-прусская война (причины и последствия) Организация работы процедурного кабинета Смысловое и механическое запоминание, их место и роль в усвоении знаний Коммуникативные барьеры и пути их преодоления Обработка изделий медицинского назначения многократного применения Образцы текста публицистического стиля Четыре типа изменения баланса Задачи с ответами для Всероссийской олимпиады по праву 
Мы поможем в написании ваших работ! ЗНАЕТЕ ЛИ ВЫ?
Влияние общества на человека
Приготовление дезинфицирующих растворов различной концентрации Практические работы по географии для 6 класса Организация работы процедурного кабинета Изменения в неживой природе осенью Уборка процедурного кабинета Сольфеджио. Все правила по сольфеджио Балочные системы. Определение реакций опор и моментов защемления |
Пояснити та схематично представити основні ефекти, місце та клітини, де утворюються такі гормони: хцк-пз, бомбезин.Стр 1 из 4Следующая ⇒
Пояснити та схематично представити основні ефекти, місце та клітини, де утворюються такі гормони: ХЦК-ПЗ, бомбезин. ХЦК-ПЗ секретується ендокриноцитами - І-клітини проксимального відділу тонкої кишки,нервові кінці дистальної частини клубової та ободової кишки.У головному мозку він відповідає за регулювання споживання їжі.ХЦК,що декретується в 12-палій та голодній кишці,представлений здебільшого ХЦК 8,ХЦК 12.ХЦК посилює дію секретину,який стимулює секрецію лужного підшлункового соку,сповільнює випорожнення шлунка,збільшує секрецію ентерокінази.Гастрин і ХЦК стимулюють секрецію глюкагону в разі перетравлення білкових страв. Бомбезин вміщує в собі 10 амінокислотних залишків.Ввін наявний у нервових волокнах блукаючого нерва,що закінчується на G-клітинах.У разі їхнього подразнення вивільняється бомбезин,що збільшує секрецію гастрину.
Пояснити та схематично представити основні ефекти, місце та клітини, де утворюються такі гормони: мотилін, вілікінін. Мотилін – поліпептид,що вміщує 22 амінокислотні залишки,декретований ентерохромафінними клітинами Мо-клітинами шлунка,тонкої та ободової кишки.У разі введення мотилі ну виникає скорочення гладких м*язів шлунка та кишки.Мотилін є головним регулятором ММК,який регулює шлунково-кишкову моторику в період між прийманнями їжі. Вілікінін – гормон,який синтезується у стінці дванадцятипалої кишки, що підвищує активність мікроворсинок у енткроцитах. Він утворюється під впливом кислого хіміку в слизовій оболонці кишки.
7.Схематично зобразити та пояснити рефлекторну регуляцію жування. Описати фази жування. Мастикаціографія.
Чорний колір – гальмівні нейрони,червоний,фіолетовий,бузковий,оранжевий – збуджуючі інтернейрони,що запукають генератор. 1 – входи для збудження мотонейронів,що піднімають нижню щелепу. 2 – що опускають нижню щелепу. 3 – гальмування мотонейронів,що що піднімають нижню щелепу. 4 – вхід збудження із вищих центрів 5 – м*яз,який піднімає нижню щелепу. 6 – м*яз,який опускає нижню щелепу. 7,8 – верхня,нижня щелепи. Акт вживання їжі умовно можна поділити на такіфази: I фаза - спокою; II фаза - введення їжі в рот; IIIфаза - орієнтовне жування; IVфаза - основна фаза жування: V фаза - формування харчової грудки та ії ковтання.
Мастикаціографія - це метод вивчення жувальних рухів нижньої щелепи. Принцип методу полягає в реєстрації коливання повітря у замкнутій системі при русі нижньої щелепи. Для запису мастикаціограми використовують гумову манжетку, трійник, затискач,капсулу Марея, кімограф, лісовий горіх. За допомогою мастикаціограми можна визначити час жувального циклу до ковтання та тривалість його окремих фаз, число жувальних рухів, величину амплітуди відкривання рота. На мастикаціограмі можна визначити також порушення жувального циклу при тій чи іншій патології, але причину, що викликала порушення циклу та його окремих фаз за допомогою цього методу виявити неможливо. 38.
39. Важливу роль у кишковому травленні відіграє жовч, яка виробляєть ся в печінці, накопичується в жовчному міхурі та надходить у просвіт дванадцятипалої кишки. Але чим вища кислотність вмісту кишечника, тим більше виділяєть
ся соку бруннерових (залози слизової дванадцятипалої кишки) і ліберK кюнових залоз (залози кишкових крипт), підшлункової залози і жовчі, що нейтралізують соляну кислоту шлункового соку і припиняють діяльність пепсину. Таким чином, існує пряма залежність між реакцією дуоденального соку і кількістю соку, виділюваного згаданими вище залозами. При цьо- му різко сповільнюються евакуаторна здатність шлунка і перехід вмісту з дванадцятипалої кишки в порожнисту.У разі зниження секреції жовчі порушаться такі процеси в тонкому кишечнику:збільшення лужної р-ції в кишечнику, за допомогою жовчі, що нейтралізує дію пепсину,зменшення бактерицидних властивостей, оскільки жовч затримує розвиток гнильних процесів у кишечнику.
40. Процеси остаточного гідролізу і всмоктування поживних речовин відбуваються на мембрані епітеліальних клітин тонкої кишки. Внутрішня поверхня кишок має вирости – мікроворсинки Мембранне травлення відбувається завдяки ферментам, фіксованим на мембранах, їхні активні центри орієнтовані на субстрат. Мембранне травлення відбувається в глибині складок мікроворсинок у стерильних умовах і тісно зв'язане з процесами всмоктування (травно-транспортний конвейєр). Порожнинне травлення здійснюється за допомогою ферментів травних секретів у порожнинах шлунково-кишкового тракту на відстані від місця утворення цих ферментів. Пристінкове чи мембранне (контактне) травлення відбувається в тонкій кишці на поверхні мікроворсинок за участю ферментів, фіксованих на клітинній мембрані. o Порожнинне травлення у людини є незначним. За типом порожнинного травлення гідролізуються високомолекулярні речовини. o Гідроліз низькомолекулярних сполук закінчується на мікроворсинках тонкого кишечника. Ферменти, фіксовані на мембранах клітин тонкого кишечнику, мають більш триваліший термін "корисної роботи" порівняно з тими ферментами, які містяться у порожнині. Особливо важливе значення мембранне травлення відіграє у розщепленні дицукрів до моноцукрів, дрібних пептидів - до амінокислот. Мембранне травлення вперше дослідив академік О.М. Уголєв. Особливостями мембранного травлення є: o висока швидкість (у десятки тисяч разів швидша за порожнинне); o здатність високомолекулярних сполук іноді всмоктуватися швидше за низькомолекулярні, що залежить від ролі транспортних білків; o висока залежність від достатності повноцінних білків у раціоні, оскільки побудоване на білках-носіях; o висока регуляторна здатність (від високої швидкості всмоктування до майже повного припинення його); o відокремленість від зони діяльності мікрофлори кишечнику.
41. Описати та пояснити механізм всмоктування продуктів гідролізу жирів. Як зміниться розщеплення жирів при зниженні секреції жовчі. НЕВЕЛИКА КІЛЬКІСТЬ ТРИГЛІЦЕРИДІВ ПЕРЕТРАВ В ШЛУНКУ ЗА УЧАСТЮ ЛІНГВАЛЬНОЇ ЛІПАЗИ.ТАКИМ СПОСОБОМ ЖИР ПЕРЕТРАВ ДО 10 %. ПРЕШИМ ЕТАПОМ ПРЕТРАВ ЖИР ПОЛЯГАЄ В ФІЗІЧНОМУ РУЙНУВАННІ КРАПЕЛЬ ЖИР НА ДРІБНІ ЧАСТИНИ ОСКІЛЬКИ ВОДОРОЗЧИННІ ФЕРМЕНТИ МОЖУТЬ ДІЯТИ ЛИШЕ НА ПОВЕРХНІ КРАПЛИНИ – ЦЕ ЕМУЛЬГУВАННЯ ЖИР. ВІН ПОЧИНАЄТЬС У ШЛУНКУ З ПЕРЕМІШУВАННЯ ЖИР З ІНШИМИ ПРОДУКТАМИ ШЛУНКОВОГО ВМІСТУЮ. ДАЛІ
ОСНОВНИЙ ЕТАП ВІДБУВАЄТЬСЯ У 12 КИШ ПІД ВПЛИВОМ ЖОВЧІ. ЛЕЦИТИН ТА ЖОВЧНІ КИСЛОТИ ЩО МІСТЯТЬСЯ В ЖОВЧІ ДОБРЕ РОЗЧИННІ ЯК У ВОДІ ТАК І В ЖИРАХ ТОМУ ЧАСТИНА ЇХ МОЛЕКУЛ РОЗЧИНЯЄТЬСЯ У ВОДІ А ІНША В ЖИР ЩО ЗМЕНШУЄ ПОВЕРХНЕВИЙ НАТЯГ ЖИР КРАП. ПАНКРЕАТИЧНИЙ ФЕРМЕНТ ЛІПАЗА МОЖЕ ДІЯТИ ЛИШЕ НА ПОВЕРХНІ ЖИР КРАП ОСКІЛЬКИ ВІН ВОДОРОЗЧИННИЙЮ ТОМУ ПОДРІБНЕННЯ ЖИРІВ В КРАЙ ВАЖЛИВЕ.КІНЦЕВІ ПРОДУКТИ РОЗПАДУ ЖИР – ЦЕ ВІЛЬНІ ЖИР КИСЛОТИ ТА 2- МОНОГЛІЦЕРИДИ ЇХ НАКОПИЧАННЯ БЛОКУЄ ПОДАЛЬШИЙ ПРОЦЕС ПРЕТРАВ ЖИР Ю ДЛЯ ЗАПОБІГАННЯ ЦЬОМУ ПРОЦЕСУ ЖОВЧН КИСЛОТ ФОРМУЮТЬ МІЦЕЛИ З ЖИР РОЗЧЧИННИМ ЯДР (ВКЛЮЧАЄ ПРОДУКТИ ПЕРЕТРАВ ЖИРІВ) І ВОДОРЧИНН ПОЛЯРНОЮ ПОВЕРХНЕЮЮ ТАКІ МІЦЕЛИ ВИКОНУЮТЬ РОЛЬ ТАКСІ ДЛЯ ПЕРЕНОСУ ПРОДУКТ РОЗЧ ЖИР ДО ОБЛЯМІВКИ ЕНТЕРОЦИТІВ. КОЛИ МІЦЕЛА ПІДХОДИТЬ ДО ЕНТЕРОЦИТА ЖИРОРОЗЧ ЯДР РОЗЧ В МЕМБРАННІ КЛІТИНИ А САМА МІЦЕЛА ЗАЛИШАЄТЬСЯ В ПРОСВІТІ КИШКИ. В КЛІТИНІ ЖИР КИСЛОТИ ТА МОГЛІЦЕРИДИ ЗАХОПЛЮЮТЬСЯ ЕНДОПЛАЗМАТИЧНИМ РЕТИКУЛОМ ТА ВИКОРИТСТОВУЮТЬСЯ ДЛЯ СИНТЕЗУ НОВИХ ТРИГЛІЦЕРИДІВ ЯКІ ЗВІЛЬНЯЮТЬСЯ ЧЕРЕЗ БАЗАЛЬНИЙ ШАР КЛІТ У ВИГЛЯДІ ХІЛОМІКРОНІВ ДАЛІ ПРОХОДЯТЬ ЧЕРЕЗ ГРУДНИЙ ЛІМФАТИЧНИЙ ПРОТОК І ПОПАДАЮТЬ В КРОВ. КОРОТКОЛАНЦЮГОВІ ЖИРНІ КИСЛОТИ Є БІЛЬШ ВОДОРОЗЧИННИМИ І ТОМУ НЕ ПОТРЕБУЮТЬ ОБОВ ЯЗКОВОГО ПЕРЕТВОРЕННЯ ЕНДОПЛАЗМАТИЧНИМ РЕТИКУЛОМОМ І ТОМУ МОЖУТЬ ВСМОКТУВАТИСЬ З БАЗАЛЬНОЇ МЕМБРОНИ ПРЯМО В КРОВ.
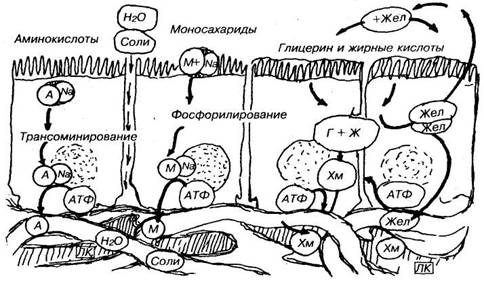
42 Представити на схемі механізм всмоктування амінокислот і моносахаридів
Всмоктування
в ротовій порожнині, в основному, всмоктуються моносахариди, деякі лікарські речовини, наприклад, нітрогліцерин; в шлунку, в основному, всмоктуються вода і алкоголь; в товстому кишечнику - вода, хлориди, жирні кислоти; в тонкому кишечнику - всі основні продукти гідролізу. У 12-палої кишки всмоктуються іони кальцію, магнію, заліза; в цій кишці і на початку тонкої кишки йде переважно всмоктування моносахаридів, більш дистально відбувається всмоктування жирних кислот, моногліцеридів, а в клубовій кишці - всмоктування білка, амінокислот. Жиророзчинні і водорозчинні вітаміни всмоктуються в дистальних ділянках тонкої кишки і в проксимальних ділянках клубової.
А - амінокислоти, М - моносахариди у сполученні з Na, Г - гліцерин, Ж - жирні кислоти - синтез уподібленних тригліцеридів у епітеліоцитах - формування Хм - хіломікронів і всмоктування в лімфу (ЛК). Жел - жовчні кислоти частково повертаються в порожнину кишечника, частково всмоктуються в кров і повертаються в печінку.
43Пояснити та схематично зобразити механізм виникнення відчуття голоду та насичення Відчуття голоду і насичення відіграють провідну роль в регуляції споживання їжі. Зовнішнім проявом голоду є поведінкова реакція пошуку їжі, направлена на ліквідацію цього відчуття. Суб' єктивні відчуття голоду при порожньому шлунку виникають періодично, в межах кожних півтори години. Вони зникають в процесі споживання їжі і не відновлюються, доки в шлунку є їжа. У формуванні відчуття голоду бере участь центральна нервова система, зокрема,гіпоталамічні структури. Експериментальним шляхом встановлено, що PDFcreated with FinePrint pdfFactory Pro trialversion www.pdffactory.com латеральні ядра гіпоталамуса відіграють роль центрів голоду, а вентро-медіальні - центрів насичення. Подразнення латеральних ядер викликає гіперфагію(підвищене споживання їжі), а їх руйнування відмову від їжі (афагію). Протилежні реакції мають мінус при впливах на центри насичення. Було висунуто ряд теорій, які намагалися пояснити механізм формування цих відчуттів і відповідних реакцій. Нині особливо багато прихільників має глюкостатична теорія, яка грунтується на результатах експериментальних досліджень, що стверджують наявність глюкорецепторів в ряді органів, в тому числі і гіпоталамусі. Саме вони сприймають зміни вмісту глюкози в крові. При внутрішньовенному введенні глюкози знижується електрична активність нейронів латерального гіпоталамуса і підвищується в вентромедіальному. Змінюється також електрична активність кори передніх відділів головного мозку, подібно тим, які реєструються у нагодованих тварин. Слід диференціювати поняття голод і апетит. Якщо голод - це стан, для ліквідації якого необхідно спожити певну кількість їжі і регулюється він гіпоталамусом, то апетит характеризується вибірковим відношенням до якості їжі, залежить від багатьох факторів і регулюється корою великих півкуль.
Пояснити та схематично представити основні ефекти, місце та клітини, де утворюються такі гормони: ХЦК-ПЗ, бомбезин. ХЦК-ПЗ секретується ендокриноцитами - І-клітини проксимального відділу тонкої кишки,нервові кінці дистальної частини клубової та ободової кишки.У головному мозку він відповідає за регулювання споживання їжі.ХЦК,що декретується в 12-палій та голодній кишці,представлений здебільшого ХЦК 8,ХЦК 12.ХЦК посилює дію секретину,який стимулює секрецію лужного підшлункового соку,сповільнює випорожнення шлунка,збільшує секрецію ентерокінази.Гастрин і ХЦК стимулюють секрецію глюкагону в разі перетравлення білкових страв. Бомбезин вміщує в собі 10 амінокислотних залишків.Ввін наявний у нервових волокнах блукаючого нерва,що закінчується на G-клітинах.У разі їхнього подразнення вивільняється бомбезин,що збільшує секрецію гастрину.
|
|||||||||
|
|
Последнее изменение этой страницы: 2016-08-26; просмотров: 242; Нарушение авторского права страницы; Мы поможем в написании вашей работы! infopedia.su Все материалы представленные на сайте исключительно с целью ознакомления читателями и не преследуют коммерческих целей или нарушение авторских прав. Обратная связь - 3.146.34.191 (0.033 с.) |

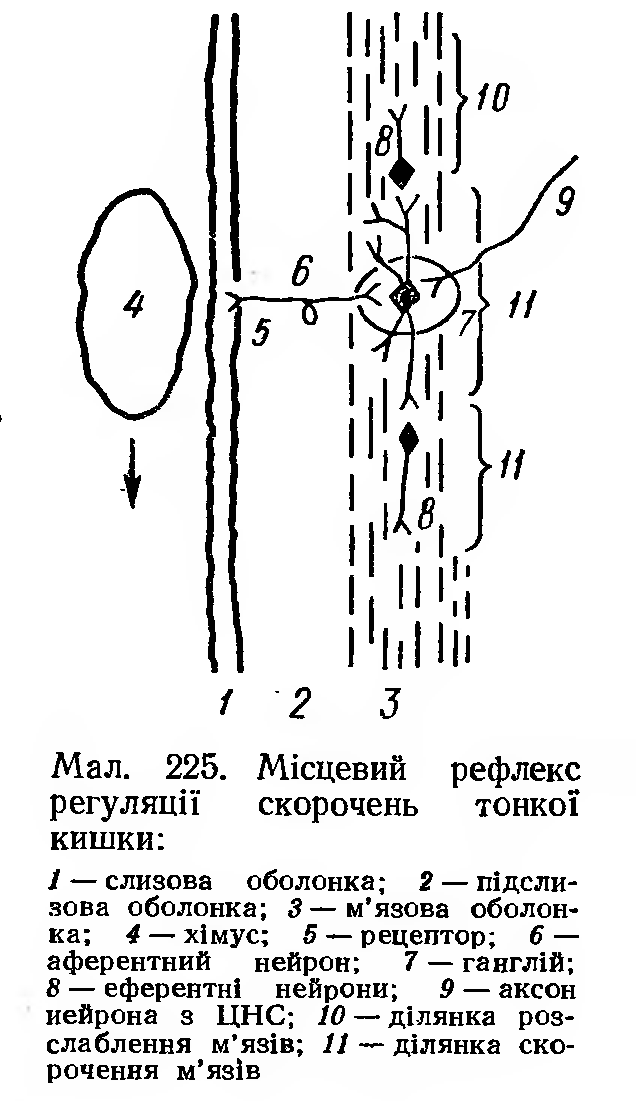
 Рефлекторну регуляцію моторної функції здійснює головним чином м’язове сплетіння у відповідь на розтягування стінки кишки хімусом.
Рефлекторну регуляцію моторної функції здійснює головним чином м’язове сплетіння у відповідь на розтягування стінки кишки хімусом.